
|
|
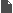 07-31-2016, 06:46 PM
07-31-2016, 06:46 PM
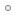
|
رقم الموضوع : [1] |
|
عضو ذهبي

 |
البستان التطوريّ {1}: تطوُّر القمح  إن ما يجمع: المعجنات، كثير من الحلويات، انواع الخبز المختلفة ...الخ، مادة سحريّة هي طحين القمح. من هنا تأتي ضرورة دراسة تطور هذه النبتة المتواضعه المسماة: قمح. سلف متواضع وشبكيّ ينتمي القمح الزراعي بشكل خاص الى نوع Triticum aestivum L ذو الاصل الحديث نسبياً، حيث يوجد منه سلالات عديدة وتتم زراعته على نطاق واسع بكل ارجاء العالم. يكون القمح، مثل الذرة، الأرز، الشعير ...الخ، ويُشكّل نبتة تنتمي الى عائلة النجيليّات، ذات دورة حياة سنويّة وتقوم بانتاج البذور. لكن، لماذا تكون ذات أهميّة كبيرة من وجهة نظر تطوريّة؟  سنابل مليئة من نوع القمح المزروع Triticum aestivum L حسناً، يتوجب البحث عن لغز هذه النبتة في جينومها. حيث تقوم كلمة واحدة بوصف طبيعتها، هي: Alopoliploide. لا تُصاب الذعر!! تعرف المؤسسة العلمية جيداً ما تقول، فتعني هذه الكلمة او المُصطلح حرفياً " مجموعات الصبغيات ( (-poli-) والغيريّة -ploidí المتعددة (alo-) "، أو بكلمات اخرى، هي عبارة عن نبتة تمتلك جينوماً مشكلاً من جينومات عديدة مختلفة. نعم كما تقرأ بالضبط!! في هذه الحالة، يمتلك جينوم نوع Triticum aestivum حجم بقيمة 16000 مليون زوج من القواعد. يكون رقم ضخم، سيما اذا اخذنا بعين الاعتبار بان جينومنا نحن يمتلك فقط 3000 مليون زوج قواعد، مُرتَّبة وفق 22 زوج من الكروموزومات الجسديّة وزوج اضافي يحدّد الجنس. من جانبه، يتكوّن جينوم نوع القمح Triticum aestivum من 21 زوج من الكروموزومات التي يمكن ان تكون مصنفه وفق: AABBDD  النمط النووي جينوم قمحنا نوع Triticum aestivum L اعتباراً من هنا يأتي ما هو ممتع فعليّاً. لدينا في الاعلى رسماً { فيه كل الكروموزومات المرتبة وفق الحجم والتشكيل } لنوع Triticum aestivum، كما يمكننا ملاحظة كل الجينوم المرتّب في ثلاث كُتل، هي: A ، B و D. توافق الكتلة D لمجموعة الكروموزومات DD المعروفة جيداً. تشكّل تلك الكتلة من الكروموزومات قسماً من جينوم القمح، وتكون هي ذاتها جينوم كامل لنوع نباتيّ آخر هو Aegilops tauschii { كما تسمى ايضاً Aegilops squarrosa }، وهي عبارة عن عشبة تنمو بشكل برّي بصيغة دغليّة { من أدغال } من اصل شرق متوسطي . سنابل قمح نوع Aegilops tauschii, Coss من جانبهما، تشكل كتلتا كروموزومات A وB مجموعة الكروموزومات AABB التي بدورها تشكّل عملياً جينوم نوع نباتيّ آخر، هو Triticum turgidum ويكون نجيليّة اخرى من اصل شرق متوسطيّ، حيث تتم زراعة بعض انواعها للآن في تلك المنطقة من العالم. هكذا وبناء على تلك المعطيات، يكون نوع Triticum aestivum في الواقع عبارة عن نوع متشكِّل من اندماج جيني وراثي لنوعين نباتيين، هما Aegilops tauschii و Triticum turgidum كنوع دغليّ ونوع " من القمح البرّي "، اللذان باتحادهما يؤصِّلان { يُعطيان الاصل } لنوع جديد بشكل كامل من القمح.  سنبلة القمح القاسي Triticum turgidum L مع ذلك، للآن لم تنتهِ القضيّة، فكما رأيتم، يتحدّد جينوم نوع Triticum turgidum بكتلتين AABB، لكن لماذا؟ حسناً، لقد اثبت العلماء عبر ابحاثهم بأننا بمواجهة جينومين أكثر. فمن جانب، لدينا مجموعة كروموزومات AA التي تنتمي للعشبة البريّة المسماة علمياً: Triticum urartu، من اقارب " القمح "، " النشويات " وغيرها. من جانب آخر، تكون مجموعة كروموزومات BB موضع جدل. حيث يوجد صعوبات بتحديد النوع الملموس الذي ينتمي له جينوم BB، توصّل بعض الباحثين لتصنيفه بنطاق مجموعة نباتية تسمى " جماعة Aegilops شعبة sitopsis "، والتي انتشرت على الصعيد العالمي عبر تشكيلة متنوعة جداً . ولم يطل المقام، الى ان تظهر دراسات جزيئية حديثة: تُشير لتحدُّر مجموعة الكروموزومات BB من نوع محدد من تلك الجماعة النباتيّة هو Aegilops speltoides، عبارة عن نبتة متوطنة في جنوب شرق اوروبا وغرب آسيا. لكن القضية لا تنتهي هنا، فاحتمال كبير أن يكون Aegilops speltoides : عبارة عن نوع قد أعطى الاصل عبر صيغة عديدة الصبغيات alopoliploidía لنوع آخر من " القمح البري " هو Triticum araraticum. ثمّة تناقض ظاهريّ، فالرؤية الشعبية للجنس Aegilops أبعد ما تكون عن القمح. يُطلق بالانكليزية عليه اسم goatgrass، والذي يعني حرفياً " عشبة الماعز " . حيث انها تمتلك فائدة وحيدة، تتمثّل باستخدامها كمرعى للقطيع. تمّ التعرّف عليها بوصفها واحدة من آباء قمحنا.  أصل وتطور الانواع المختلفة من القمح بالتالي ذات الصيغة السالفة الذكر، فنوع Triticum turgidum بدوره نوع آخر يأتي جينومه من اتحاد جينومين لنوعين مستقلين، يمكن ان يُعثر عليهما كاعشاب برية كما اعشاب مزروعة موطنها شرق المتوسط . التطور والتآلف اللذان اعطا الاصل لقمحنا ربما لا يمكن تفادي السؤال: كيف يمكن ان يحدث هذا؟ كمثالن نعرف بأننا لو نقم بتهجين حمار وفرس سنحصل على بغل، وهو حيوان مصاب بالعقم كليّاً. يتأسّس الحلّ على واقع أنّ في مجموعات نباتية كثيرة، لا تعمل هذه الصيغة { وهذا ايضا سبب صعوبة تصنيفات نباتية لجماعات نباتية ملموسة في كثير من الاحيان }. حيث انه في هذه الحالة، يحدث الآتي: اولاً: تشكل النجيليات نباتات تمتلك تكاثر هجينيّ بواسطة الرياح، حيث يكون غريباً أنّ غبار طلع لنوع يمكن ان يصل لزهور انثوية لنوع آخر.  ازهار نوع القمح ، Triticum aestivum, L ، لاحظوا تموضع الاسدية { السداة هي العضو الذكري في الزهرة } التي تسهل نثر غبار الطلع بواسطة الرياح ثانياً: لسبب ما، تكون النباتات أكثر سلاسة وراثياً جينياً من الحيوانات، بصيغة تُمكِّنها من اعطاء ظواهر متعددة المجموعات الصبغية وحتى لتعدد مجموعات صبغية بين انواع مختلفة. حيث نرى بأن 30 نوع مُكتشف بين جنسي Triticum و Aegilop: يكون 75% منها على الاقل معروفة بكونها alotetraploides { ناتجة عن اتحاد نوعين } أو alohexaploides { ناتجة عن اتحاد 3 انواع }. ثالثاً: لا تكون تلك النباتات هجائن بسيطة، حيث يجب أن يمتلك الهجين الصرف نصف جينوم من نوع ونصف آخر من نوع آخر، وكما من المعتاد حصوله في الحيوانات، إما تكون مُصابة بالعقم أو تمتلك خصوبة تالفة. تكون هذه الحالة مختلفة، هي ظواهر تعدّد للمجموعات الصبغية { تتضاعف المادة الوراثية لاحد تلك الهجائن }، وبهذا نمتلك جينومات كاملة وظيفيّة لانواع محددة. حيث يمتلك الافراد الناتجين بالعموم اكبر قدر من الخصوبة والقوة الفيزيائية. في الواقع، يمكن ان تكون ظاهرة تعدد المجموعات الصبغية الغيريّة عامة بصيغة أكبر مما هو مُنتظر، وتكون قادرة على المساهمة في نشوء سريع للانواع بين النباتات، وهذا يتسبب بتغيرات هامة في تنظيم ونشاط المادة الوراثية. بكلمات اخرى، تشكّل قوّة فعّالة محرّكة للتطور. لا تكون قواعد تعدد المجموعات الصبغية صعبة على الفهم. ففي الجدول التالي يتم توضيحها بشكل جيد: جدول يلخص تعدد أو تغاير المجموعات الصبغية alopoliploidía لنفترض وجود نوعين، نوع A ونوع B ، كلاهما متعدد الصبغيات، ما يعني، يمتلك كل واحد منهما مجموعه مضاعفة من كروموزومات (2n) في داخل خلاياه، والذي يكون في النوع A هو: AA، وفي النوع B سيكون: BB . خلال التكاثر الجنسي، يُنتج خلايا متخصصة لاجل التناسب مع مجموعة وحيدة من الكروموزومات (n) لكل واحدة منها، تستقبل تلك الخلايا اسم الامشاج { عند البشر هم البويضات والحيوانات المنوية }، بهذا يمتلك النوع A امشاج A والنوع B امشاج B . ففي الالقاح العادي بين اعضاء من ذات النوع، يتحد مشيج ذكري (n) مع مشيج انثوي (n): مُعطياً لاقحة (2n). لكن خلال عملية تهجين، ما يحصل هو اتحاد مشيج نوع (A) مع مشيج نوع آخر (B) ما يؤدي لظهور لاقحة هجينة تمتلك مجموعة وحيدة من الكروموزومات لكل نوع (AB). تلك الهجائن عند الحيوانات، من النادر ان يكون لها مستقبل. لكن عند النباتات، يكون شائعاً بلوغها عمر التكاثر، على الرغم من ان اغلبية المرات ستكون مصابة بالعقم، او تقريباً مصابة به. مع هذا، يمكننا الآن الالتفات الى ظاهرة معتادة في الطبيعة هي الطفرات. لكن بحالتنا لا يتم الحديث عن أيّ طفرة، بل الحديث عن الطفرة: الآلة الجينية الوراثية الاكثر بهيميّة التي يمكن تخيلها. تضاعف كليّ وشامل للمادة الوراثية. يمكن ان تحدث عبر التلوث الكيميائي بمركبات مثل colchicina، في اخطاء خلال نشوء انفصال الكروموزومات في الانقسام الاختزاليّ meiosis ، أو بسبب ثغرات في الانقسام الخليوي. ستكون النتيجة امشاج بترتيب جيني 2n أو في حالة الهجائن مع ترتيب جيني AB. كيف يؤصّل هذا لمتعدد أو متغاير المجموعه الصبغية؟ حسناً، تقتضي الخطوة التالية واحدة من " اللاترجيحات الكونيّة "، تستلزم حادث تخصيب بين امشاج ثنائية المجموعة الصبغية. فهكذا مشيج ثنائي (2n) + مشيج ثنائي آخر (2n) سيعطي المجال لظهور = لاقحة متعددة المجموعات الصبغية (4n). في حالة الهجائن، ستكون النتيجة عبارة عن كائن: AB + AB = AABB، ويكون هؤلاء الافراد بعكس الهجين الذي أصَّلهم { أصّل، يؤصِّلُ .. يُعطي الأصل لكائن حيّ أو لشيء ..الخ } انواع فعّالة وذات خصوبة جيّدة. بكلمات اخرى، انواع كاملة، جديدة، حرّة ومستقلة. لهذا، لا يكون قمحنا سوى احد الامثلة الاكثر وضوحاً عن التطور، الذي يمكننا العثور عليه كذلك من خلال التغيرات المورفولوجية والوراثية الهامة. من جانب آخر، على الرغم من كون نموذج نشوء انواعه غير مألوف كليّاً في المملكة الحيوانية، فإنّ تعدد المجموعات الصبغية يشكّل ظاهرة عادية في المملكة النباتيّة. علم الوراثة والتدجين أين ومتى تمّ اعطاء الاصل لقمحنا Triticum aestivum؟ على الرغم من ان الامر للآن موضع جدل، لكن يتفق كثير من الباحثين: على ان هذا قد حدث في منطقة شرق المتوسط، وقد حصل منذ 11000 عام حيث حصلت موجات ذوبان جليدية بآخر عصر الجليد، عندها انتشرت انواع مختلفة من البقول البرية لتصل الى منطقة الهلال الخصيب Creciente Fértil { المنطقة الممتدة بوقتنا الحالي لتشمل فلسطين، الاردن، لبنان، الجمهورية السورية والعراق } . استقرت بهذه المنطقة عدة انواع من القمح البري: Triticum monococcum, T. urartu, T. boeoticum, T. dicoccoides, Aegilops tauschii). ومن الشعير البري (Hordeum spontaneum) ومن الشوفان البري (Secale vavilovii) منطقة الهلال الخصيب كان احد اهم خيوط التاريخ البشريّ: التحوّل من مجتمعات التقاط الثمار والصيد المتنقلة الى جماعات مستقرة متخصصة بالزراعة. ففي غرب آسيا، يُسجّل تحول كهذا عبر ثلاث انواع من النجيليات البرية، والتي يُعتبر الانسان من وقتها المتسبب بنهبها المستمر: نوع من القمح البري la escanda، القمح والشعير. تسجلكثير من الدراسات الجينية بأنّ اصل بذرة تدجين تلك النباتات، أتت منمنطقة دياربكر الواقعة بجنوب شرق تركيا الحالية. من هنا انطلقت بداية زراعة " القمح البري الاصغر " عبارة عن نوع قمح (Triticum monococcum) ذو جينوم نموذج AA ومن اقارب نوع Triticum urartu السلف لقمحنا الحالي. كما ان تلك المنطقة عرفت البذرة المزروعة من القمح نوع T. turgidum ssp. Dicoccoides المعروف بيومنا هذا تحت اسم " النشويّ البريّ ".  سنابل نوع القمح البري الاصغرTriticum monococcum في الاعلى سنبلة لنوع قمح مزروع، وفي الاسفل سنبلة قمح برّي وتُعتبر فلسطين من اوائل الاماكن التي استعمل فيها القمح، حيث يمكننا الحديث عن باقي " النشويات البرية "، مثل: Triticum turgidum ssp. Dicoccoides { تُعتبر في زمن آخر كنوع مستقل هو Triticum dicoccoides } كسلف مُباشر { او مباشر تقريبياً } لقمحنا الراهن. على الرغم من ان الاستعمال لا يعني التدجين، فإنّ اوائل الطبقات التي ظهرت فيها تلك الانواع تعود الى 19000 قبل الميلاد. فكما قلنا، بمجرد حصول تكييفها للزراعة، اعطت المجال لولادة الزراعة. يظهر تدجين هذا النوع في مناطق الهلال الخصيب اعتباراً من القرن الحادي عشر قبل الميلاد، بينما يعود ظهورها البرّي الى 13000 عام قبل الميلاد في الجنوب الشرقي لتركيا الحالية. مقارنة بين النوع الخشن Triticum turgidum ssp. durum الى يسار الصورة ونوع قمحنا الحديث Triticum aestivum إلى اليمين لقد رأينا للآن بأنّ الظاهرة الابرز لفهم تطور قمحنا، قد كانت: تعددية المجموعات الصبغيّة poliploidía، مع ذلك، يُعتبر التدجين ايضاً قد لعب ورقة هامة. فبخلاف قمحنا الحديث، يكون النشويّ البريّ عبارة عن نبات ذو بذور صغيرة الحجم، ويكون انتاجها بكميات قليلة، اضافة لكونها محمية بواسطة قشرة. ليس هذا فحسب، مثلما يكون كثير من النجيليات فإنّ نوع Triticum turgidum ssp. Dicoccoides عبارة عن نبات وسط نشره الطبيعي الرياح والحيوانات. لاجل هذا يحتوي على سنابل متفتحة " هشّة "، بحيث تنكسر تلك السنابل بسهولة، بمجرد نضوج البذور: محررة سنيبلاتها بصيغة فرديّة. تشكّل تلك السنيبلات وحدة نشر هذا النبات، كوحدة تشكيل ازهار اساسية في النجيليات، ومن ثمّ تستقبل البذرة. هذا لا يحصل هكذا في قمحنا، في الواقع سيكون عبارة عن مشكلة. هل بامكانكم تخيّل سنبلة " دون حبوب " وقت الحصاد؟ بالتالي، يكون الاكثر اهمية بالنسبة لنا هو سنبلة تحتفظ ببذورها بكل لحظة. مع ذلك، في الطبقات الاكثر قدماً، الموجودة في الجنوب الشرقي من تركيا الحالية وشمال سورية الحالية، والتي تعود الى 10200 عام، تنتمي كل البقايا الاثرية عملياً لسنيبلات متفتحة " هشّة ". لكن الوضع قد تغيّر. ففي الطبقات التي تعود الى 7500 عام، فنحن امام وضع فيه 30% من السنيبلات من النوع المُنغلق او المُطبق " الغير هشّة ". وبمرور 1000 عام، امتلك اكثر من 60% من تلك السنيبلات خصائص كتلك. تكون السنبلة المنغلقة او المطبقة: تلك التي تحتفظ بكامل بذورها عند النضوج، فقد أضحت عملياً السمة الثابتة في القمح.  السنبلة الحديثة المنفتحة { او " الهشّة " } للنشويّ البريّ A ، وسنيبلة منها B . تفاصيل لمقطع من ذات السنيبلة C قمح مدجّن منغلق D. وتفصيل من عينة عشوائية لسنيبلة منه E ، الرسم البيانيّ F يوضح تناوب عدد من الاحفوريات ذات الصلة لانواع هشّة منفتحة ولانواع منغلقة قاسية.طبقات موقع الأسود Aswad وطبقات موقع الرماد Ramad تنتمي لحالات من الشعير في الوقت الذي عادت فيه السنابل منغلقة، ايضاً كانت ميزات اخرى قيد الانتقاء. سنابل منغلقة مع بذور اكثر صلابة: هو ما يمكننا رؤيته في النشويّ المزروع، الذي يسميه بعض الباحثين Triticum turgidum ssp. Dicoccon. ملمح آخر يميزه عن النشويات: هو حضور بذور مغطاة ومحمية بقشرة. وتلك ميزة مضبوطة جينياً بما يسمى loci Tg (tenacious glume) وعناصر del locus Q. لكن تلك الميزة السلفيّة { من السَلَفْ } ايضاً كانت في مهب الاختفاء. وبسرعة تظهر بذور حرّة من القشرة في طبقات تعود الى 8500 عام. شكّلت تلك الصيغ النيوليتية السلف المُباشر لقمحنا الحديث كما " للقمح القاسي " الذي ما يزال يُزرع في شرق المتوسط وشمال افريقيا، والمعروف تحت اسم Triticum turgidum ssp. Durum، عبارة عن نبات قد ورث كل الصفات الجيدة { من وجهة نظر زراعية } التي ظهرت في النشويّ، الى ما تمّت اضافته عبر حضور حبّة عارية، حرّة من القشرة، الامر الذي يسهل استعمالها ومعالجتها.  التعاقب التطوري لنوع قمحنا الحديث، فمن اليسار الى اليمين نجد: النشويّ البريّ A ، النشويّ المزروع B ، القمح القاسي C والقمح الحديث D ، الاحرف المتموضعة في الاسفل تمثل جينات تلك الانواع بالنهاية، بلغ تدجين القمح ذروته بظهور نوع Triticum aestivum. الذي كان نتاج تعدد تنوعات نوع Triticum turgidum من بذور عارية وسنابل منغلقة في الغالب، ومن خلال " عشبة الماعز " تلك المسماة Aegilops tauschii. الامر الذي يُعطي الاصل لقمحنا الحديث، كنوع خاص حديث جداً. على الرغم من ان عمره لا يكون متناسقاً بكل حساباته، فإنّ الحساب على قاعدة ساعاته الجزيئية اجراه بعض الباحثين: حددوا عمره بحدود 9500 قبل الميلاد، بينما يُرجعه باحثون آخرون لزمن ابكر لحوالي 6000 عام قبل الميلاد، فيربطونه ببدء الزراعة جنوب غرب بحر قزوين: المكان الذي تنتشر فيه " عشبة الماعز " تلك بصورة طبيعية. تمّ تبرير نجاح نوع القمح Triticum aestivum تقليدياً بمواجهة " القمح القاسي " عبر الاستعانة بصلابته العالية، الامر الذي شكّل افتراضاً اعرجاً! حيث يُنتج " القمح القاسي " بذور اكبر حجماً ويمتلك قدرة على انتاج مشابه بشروط نمو مفضلة. من وقت قريب جداً، طرح باحثون آخرون مقارنة لانواع قمح : tetraploides , Triticum aestivum الامر الذي يوفّر لدونة عالية بمراحل مختلفة وشروط الاستبراد { معالجة البذور بالبرد بُغية تسريع انتاشها او انباتها وتسريع نموّ النبات وازهاره .. قاموس المورد اسباني عربي }، سامحة له بالنمو والانتاج تحت تأثير ظروف مُناخية مختلفة، تطوير مقاومة للملوحة، انخفاض قيمة pH لحضور الالمنيوم وللتبريد، مقاومة عالية للاوبئة والامراض، كذلك امتلاك قدرة عالية لاجل تحضير نماذج مختلفة من المنتجات الغذائية.  تطور القمح. لاحظوا التغيرات في شكل السنبلة  الموضوع الأصل بالقسم الأجنبي تعليق فينيق تتكون سلسلة البستان التطوري من عشر مواضيع، وتتناول تطوُّر أنواع نباتيّة محددة، حيث تبدأ السلسلة بتطور القمح، تتميز هذه السلسلة بغناها بالمعلومة والتوثيق. سأنشر المواضيع على مداخلات متتابعة هنا. أشكر أيّ تصويب أو إضافة   |
|
|
|

|
|
07-31-2016, 07:01 PM
|
رقم الموضوع : [2] |
|
عضو ذهبي
|
البُستان التطوريّ {2}: تطوّر العنب مشروب الآلهة اليونانيّة تستمر مشروبات ديونيسوس وباخوس بفضل تخمير عصير العنب أو ثمرة الدالية. يكون الاسم العلمي لهذا النبات Vitis vinifera subsp. Vinífera وينتمي الى عائلة Vitaceae التي تضمّ اكثر من 1000 نوع حيّ من النباتات التي تعيش بمناخ مداري أو تحت مداري، والتي تنمو كعرائش. بدأ تدجين هذا النبات منذ 8000 عام، ومن وقتها قام الانسان بتوجيه تطوره عبر الانتقاء الصناعي. مع هذا، تعود بدايات ظهور هذا النبات الى زمن أبعد بكثير مما تتسع له ذاكرة النوع البشريّ.  نوع عنب Vitis vinifera ssp بوضع صالح للقطاف عرض أوّلي بمقارنة نوع Vitis vinífera بانواع اخرى، نراه يمتلك جينوماً متواضعاً نسبياً، حيث يمتلك بين 475 و500 مليون من ازواج القواعد فقط، ودون الذهاب بعيداً، يمتلك جينومنا البشري 3000 مليون زوج قواعد، ويمتلك جينوم القمح 16000 مليون زوج قواعد. في نوع العنب ذاك: يتكون الجينوم من 19 زوج من الكروموزومات وبشكل مضاعف، حيث يكون كل زوج من الكروموزومات لوحده، وبذلك لا يكون لدينا حالة تعدّد للمجموعات الصبغية و تعدد للمجموعات الصبغية الغيريّة poliploidías, alopoliploidíasوغيرها من الحيل الجينية ..  تمثيل الاجزاء المختلفة بجسم شجرة العنب نوع Vitis vinifera ssp. Vinífera سجّل العام 2007 علامة فارقة بالنسبة للدارسين، للمُنتجين، للتجار، لعشّاق النبيذ. حيث تمّ انجاز سَلْسَلة جينوم نوع عنب Vitis vinifera ssp. Vinífera ويكون الجينوم الرابع بتسلسله لنبات زهريّ، والجينوم الثاني لنبات خشبيّ والجينوم الاول لنبات ثمريّ. قاعدتين نباتيتين من ثلاث، لا يكون الامر سيئاً. طابق تحليل جينوم نوع Vitis vinífera ما كان معلوماً: بانه لا وجود لاي ظاهرة حديثة لتعدد المجموعات الصبغية. ما قوله، خلال الملايين الاخيرة من الاعوام، ولا اي سلف من اسلاف نوع Vitis vinífera قد وُلِدَ اعتباراً من تضاعف جينوم نوع سابق. لكن قلنا " حديثاً " و " الملايين الاخيرة من الاعوام ". المفاجأة: حصل هذا عندما " رجع " علماء الوراثة أبعد بكثير من ذاك الزمن. لقد رأينا سابقاً بأنّ كلا تعدد المجموعات الصبغية الغيريّة وتعدد المجموعات الصبغية، لديهما القدرة على توليد انواع جديدة خلال زوج من الاجيال. تشكل تلك الحالة عودة التفافية اكثر. يكون جينوم الدالية هو نموذج paleohexaploide وهو عبارة عن مصطلح مركّب يعني حرفياً " مجموعة كروموزومات (-ploidía) سداسيّة (-hexa-) وقديمة جداً (paleo-) ". يمكننا قوله بطريقة اخرى: عبارة عن نبات، ظهرت اسلافه البعيدة بفضل تثليث الجينوم لنوع سابق. " العلم الاحاثيّ الجيني Paleogenética ". عودة نحو الماضي من الطبيعي ان يقوم اي شخص بالتشكيك حول هذه النماذج من التأكيدات. لا نمتلك اي جهاز زمن يدوي، فنحن نعرف او ربما نعرف، فيما لو توجد قاعدة في عالم العلم، ستتمثّل بترك الماضي لندبات أو لآثار يمكن دراستها في الوقت الحاضر. ولن تكون المادة الوراثية الجينية شيء اقلّ بهذا الصدد. قَبِلَ علماء الوراثة او الجينات بهذا المضمار الاتجاهات العامة لظواهر تعدد المجموعات الصبغية في النباتات، ولاجل ذلك ركّزوا بحثهم في مناطق متناددة { في علم الأحياء التطوري، التنادد هو أي تشابه بين مميزاتعضيات بسبب انحدارهم من جد مشترك. أكواد الدنا ذوو التسلسلات المتشابهة يـُفترض أن لهم سلف مشترك. الكروموسومات المتناددة هي كروموسومات لها نفس الجينات ونفس التسلسلات. ترجع أهمية التنادد في التكاثر إلى أن أزواج الكروموسومات المتناددة تصطف معاً أثناء الانقسام الاختزالي. منقول يُرجى التدقيق } مناطق التنادد Parálogas من الكروموزومات { لن ننسّ بأن الكروموزوم Cromosoma يعني بالعربية الصبغيّة : التي هي قطعه من الخيط الصبغي الذي يظهر في نواة الخلية اثناء انقسام تلك النواة. ويكون عدد الصبغيات واحداً في كل نوع نباتيّ .. قاموس المورد اسباني عربي }.  جدول يبين منطقة تنادد في الاعلى كروموزوم اصلي، في الوسط حصول تضاعف في الجين، في الاسفل تراكم الطفرات وتشكّل منطقة تنادد تكونا منطقتا تنادد " منطقتان غنيتان في الجينات المتشابهة، حاضرتان ضمن ذات الجينوم ". تتشكلان بفضل تضاعف داخلي للجينات والتراكم اللاحق للطفرات. بفضل تآلف كلا الحادثتان، تشكل منطقتا تنادد متميزتان بذاتهما، ما يشكل طابع هويّة ذاتيّة. بحيث انه تتحدر منطقة تنادد A فقط من منطقة تنادد اخرى A سلف. لكن، فيما لو تكن تلك " المناطق الغنية بجينات متشابهة " والحاضرة " ضمن ذات الجينوم "، والتي كانت " في جينومات مختلفة " فإننا نتحدث عن مناطق منضبطة regiones ortólogas. تكمن منطقية اهميتها نسبة لمناطق التنادد regiones parálogas بكونها هي ذاتها، الامر الوحيد المُختلف هو سياقها. حسناً، ستقولون " لكن هذا لا يقول لنا شيء عن كيفية السفر { أو الرجوع } في الزمن ". عندكم حقّ، يتوجب علينا التقدّم خطوة أكثر. ثبّتوا نظركم في الرسم البياني التالي:  رسم توضيحي للفارق بين مناطق التنادد والمناطق المنضبطة كما قلنا سابقاً، بسبب تعقيدها، سواء مناطق التنادد أو المناطق المنضبطة، فهي تعبر عن طابع هوية ذاتية، الامر الذي يستلزم حضور صيغة من القرابة. وفق هذا الامر، فيما لو نعثر على منطقة منضبطة في نوع A وذات المنطقة المنضبطة في نوع B ، فهذا يعني امتلاك كلا النوعين A و B لسلف مُشترك، قد حمل تلك المنطقة المُنضبطة ايضاً. وما الذي يحدث فيما لو نعثر على منطقة تنادد في الكروموزوم A وذات منطقة التنادد في الكروموزوم B ؟ بحسب الاستنتاج السابق، ستكون كروموزومات A و B متصاهرة { تمتلك قرابة }، مع ذلك يوجد فارق نسبة للوضع الداخليّ، فقد كانت كروموزومات A و B متصاهرة فقط، بل انها كانت هي ذاتها. أيّ نموذج من الظواهر نعرف بكونه قادر على اعطاء الاصل لزوج من الكروموزومات المتساوية بذات الفرد؟ فعلياً، لدينا نموذج تعدد المجموعات الصبغية poliploidías، وبهذا نمتلك الندبة { الأثر } التي نبحث عنها لاجل انجاز سفرنا عبر الزمن. من المفترض وجود " عقبة " في الاستدلال السابق. حيث يوجد في الكروموزومات عناصر مثل الينقولات transposones { الينقول (بالإنجليزية: Transposon) وهي سلاسل من الدنا DNA يمكنها التحرك اٍلى مواقع مختلفة داخل الجينوم الخلوي لخلية بعينها، تسمى العملية بالاٍنتقال. يمكن خلال الاٍنتقال أن تحدث طفرات أو تغير جزء من الدنا في الجينوم. هناك من يسمي الينقولات بالجينات القافزة، و تعتبر مثالا عن العناصر الوراثية المتحركة التي اٍكتشفتها بربرة مكلنتوك في بداية سيرتها العلمية و التي نالت بفضلها جائزة نوبل لعام 1983... منقول يُرجى التدقيق } والمعروفة تحت اسم " عناصر وثّابة ": عبارة عن تعاقبات من الدي ان اي، تنسخ بسهولة كبيرة وتقفز من جانب لآخر من الجينوم، داخلة باي جانب تقريباً مع قدرة كليّة على الافلات، مما يمكنها من نقل مقاطع مستقيمة من الجينات في طريقها. لهذا تمتلك القدرة ايضاً على نسخ ونقل مناطق تنادد. اذاً، كيف يمكن حلّ هذه المشكلة؟ الجواب آتٍ، لا تستعجلوا!! لندخل الآن بعمق في موضوعنا. ما الذي رآه علماء الوراثة عندما درسوا جينوم الدالية المزروعة؟ تأمّلوا الصورة التالية، وبالتفاصيل بالاجمال:  ترتيب مناطق التنادد في مختلف الكروموزومات تُشير الالوان لمناطق التنادد بين الكروموزومات الاكثر تمثيلاً في الدراسة مثلما ترون، يحتوي كل كروموزوم على الاقلّ زوج من مناطق التنادد regiones parálogas . وكل زوج من تلك المناطق يتمثّل حصرياً بثلاث كروموزومات مختلفة، ولا واحد اكثر ولا واحد اقلّ { إلا بحالات استثنائيّة }. انه كرؤية مجموعة n كروموزوم مضروبة ب 3. ومن جانب آخر، من الصعب تفسير هذه الدقّة عبر فعل صرف متمثل " بعناصر وثّابة ". { وثّابة او نطّاطة }.  مقارنة بين النواة الخلويّة البشرية H والنواة الخلويّة للشمبانزي C ، لاحظوا الارتباط الجيد بين الاشرطة. لهذا لا يكون صعباً الاستنتاج بناء على ذلك بأنّ جينوم الدالية للآن يحتفظ بندبات { المقصود بآثار قديمة }، حيث كان في الماضي عبارة عن جينوم متعدد مجموعات الصبغيات قديم جداً، تحديداً نوع hexaploide، بوصفه ثمرة تثليث جينوم لسلف اقدم اكثر. { يعني hexaploide حرفياً كائن ذو 6 جينومات ومن هنا نفهم معنى التثليث: 2.3 .. اقتضى التنويه فينيق }. إلا أنه يتبقى بعض الاشياء المحتاجة للتوضيح. مثل: كيف يُعرف بكونه قديم؟ وفيما لو يكن جينوم ثلاثيّ { مُضاعف ثلاث مرات }، لماذا يمتلك 19 كروموزوم وليس 18 أو 21 { من مضاعفات العدد 3 }؟ حسناً، يمكن اكتشاف عمره بعدّة طرق. الطريقة الاولى عبر نسبة التعديل الكبيرة فيه، فكما رأيتم في الصورة بالاعلى، تكون مناطق التنادد متوزعة في اماكن مختلفة من الكروموزومات، يكون بعضها معدلاً حتى، وحتى امتلاكه 19 وليس 18 او 21 زوج من الكروموزومات هو عبارة عن دليل اضافي على اقدميته. حيث لا غرابة بالامر، يمكن للكروموزومات ان تتغير كثيراً بطول زمن التطور. تكون قادرة على تبادل الجينات، الدوران 180 درجة، الانقسام، الاندماج .. الخ. دون الذهاب بعيداً اكثر، تكون كروموزماتنا 2 متطابقة مع كروموزومين للشمبانزي، لكن تكون مندمجة. نعرف هذا بفضل الارتباط التام للكروموزومين الحرين للشمبانزي وكروموزماتنا 2. دليل آخر اضافي، تُولد أقدميّة نوع hexaploidía من مقارنة جينوم الدالية مع جينومات نباتات اخرى تمّ سلسلتها من وقت قريب، مثل: حور كاليفورنيا Populus trichocharpa thalianaarabidopsis - Arabidopsis { وهونوع قريب من الفصيلة الصليبية أو الكرنبية أو الخردلية هي إحدى أهم الفصائلالنباتية تنتمي إلى رتبةالكرنبيات. يبلغ عدد الأنواع التي تتبع هذه الفصيلة حوالي 3000 نوع معظمها أعشابحولية أو معمرة. تضم هذه الفصيلة الكثير من محاصيل الخضراوات مثل الملفوفوالفجلواللفتوالقنبيطوالقنبيط الأخضروالكرنبوالجرجيرولفت الشلج .. منقول يُرجى التدقيق }،والأرز (Oryza sativa) . فرضية الانتقال تُثبت فيما لو تكن مناطق التنادد محفوظة جيداً: فعند مقارنة تعاقباتها مع تعاقبات تلك النباتات، سيتوجب اعطائها للنتائج التالية: أولاً: مع نباتات نوع hexaploides قديمة، سيُوجب كون الاصطفافات بين المناطق، السبب بظهور طبقة في الدالية مقابل كل منطقة في النباتات الاخرى { نموذج 1:1 }. ثانياً: مع نباتات غير نوع hexaploide القديمة، سيُوجب كون الاصطفافات بين المناطق، السبب بظهور 3 طبقات في الدالية مقابل كل منطقة في النباتات الاخرى { نموذج 1:3 }.  مقارنة تعاقبات الدالية مع: a حور كاليفورنيا، لاحظ الكتلتين والاصطفاف نموذج {1:1} في كل كتلة. مع bنوعArabidopsis لاحظ الكتل الاربعة والاصطفاف، من النموذج { 1:1 } تقريباً. مع cالارز، لاحظ المزيج الناتج عن الاصطفاف { نموذج 1:3}. وما الذي رأيتموه؟ انظروا للصورة فوق: a- مع نوع حور كاليفورنيا، انشطرت الاصطفافات الى كتلتين كبيرتين، أمر مُفسّر بشكل كليّ، فمن المعروف بأنّ نوع Populus trichocharpa قد ورث جينوماً مضاعفاً. والافضل، كانت الاصطفافات في كل كتلة من النموذج 1:1. b- مع نوع Arabidopsis thaliana، بيّنت الاصطفافات 4 كُتل كبيرة، ايضا سهلة التفسيرن حيث انه معروف بأنّ نوع Arabidopsis thaliana قد ورث حادثين ضمن سلسلة من التضاعف الجينومي بطول نَسَبِهِ الحديث. وايضاً في كل كتلة: ظهرت اصطفافات من نموذج 1:1. c- مع الارز كانت القصة مختلفة. فقد التقت اغلبية الاصطفافات بذات المكان، ما يعني اعطائها المجال لظهور النموذج 1:3. والذي لم يكن بالتالي من نوع hexaploide القديم. هذا يُشير لأنّ نوع hexaploide سداسي الجينومات السلف لداليتنا، قد ظهر فقط في الفترة الواقعة بين انشطار النباتات الى monocotiledóneas { صف احاديات الفلقة بالعربية } و { ثنائيات الفلقة } dicotiledóneas { بين 130 الى 240 مليون عام } وظهور السلف المُشترك لحور كاليفورنيا نوع arabidopsis والدالية يعود الى الفترة الممتدة بين { 106 الى 112 مليون عام }. دعمت دراسات لاحقة حول كاسيات البذور النموذج الاخير.  نتائج جينية وراثية لظهور hexaploide الاصلي. تمثل النجوم في الرسم حوادث التضاعف الجيني. التدجين وتبعاته ها نحن نعرف بأن حوادث قد حدثت منذ زمن قصيّ، زمن اسلاف الدالية { الكرمة، شجرة العنب .. كما تريدون }. لكن ما الذي حصل في العشرة آلاف عام الاخيرة؟ نعثر في هذه الحقبة الحديثة على السلف المُباشر والبريّ للدالية المزروعة، اسمه العلمي هو: Vitis vinifera ssp. Sylvestris. حيث لا يكون نوع مُختلف عن نوع Vitis vinifera ssp. Vinífera، بل يكون كلاهما : نوعان فرعيان.  مثال من نوع دالية Vitis vinifera ssp. Sylvestris وما الذي يعنيه: نوع فرعيّ؟ بكلمات قليلة، عبارة عن جماعات من نوع حيّ معين اكتسب مستوى ملموس من الاستقلال الوراثي، وبهذه الصيغة، تكون جماعات في منتصف طريق التحوّل الى نوع حيّ مستقل. في حالتنا، يكون نوع Vitis vinífera عبارة عن " نوع في منتصف الطريق لينقسم الى نوعين " نوعين مختلفين من الدوالي. بالتأكيد، تكون الفروقات بين النوع البريّ والمزروع واضحة، كما في الشكل التالي:  الفرق بين الدالية المزروعة { يسار الصورة } والدالية البرية { يمين الصورة } كل هذه الفروقات او التغيرات، الظاهرة للبعض والغامضة للبعض الآخر: تشكّل مؤثرات لبضع آلاف من السنين من التدجين والانتقاء البشريّ لها. يكون نوع Vitis vinifera ssp. Sylvestris بوقتنا الراهن نادر جداً. فقط تمّ العثور عليه في المانيا، النمسا، اسبانيا، فرنسا، ايطاليا، رومانيا، سويسرا وبلدان اوروبية اخرى، وباكثر الاحوال عُثرَ على عينات قليلة من المعتاد تهجينها مع النوع الفرعي المزروع. ايضاً يوجد نماذج في جورجيا، ايران وتركيا، والتي من المفترض ان تكون المناطق الاساسية لاصل هذا النوع من النباتات.  طقس شرب النبيذ في احتفالات اليونان القديم فعلياً، تعود اوائل الادلة على بذور مدجنة وانتاج للنبيذ، لحوالي 8000 عام قبل تاريخنا الحاضر في منطقة جورجيا، ايران وتركيا. انتشرت زراعة هذا النبات سريعاً فوصل الى مصر، الهلال الخصيب واوروبا المتوسطية، قبل نهاية الالف الخامسة قبل تاريخنا الحاضر. كيف لا، وقد كان اليونانيون بارعون في صناعة النبيذ منذ 6500 عام قبل تاريخنا الحاضر ومعهم ظهر الاله ديونيزوس، احد افضل الآلهة التي صنعها العقل البشريّ. يتحدث بعض الباحثين عن حادثي تدجين: الاول في الهلال الخصيب، والثاني في اوروبا الغربية. ففي اواخر قرون 3000 قبل تاريخنا الحاضر، بلغت شعبية النبيذ اوجها بوصولها الى الصين واليابان. فتحوّل النبيذ من وقتها لصديق كبير للانسان. بلغت شعبيّة النبيذ في وقتنا الراهن اقصى مدى، حيث يوجد علم متخصص بدراسة تحضير النبيذ، هو: enología { علم الخمور والأنبذة: (بالإنجليزية: Enology) هو العلم الذي يختص بدراسة جميع الجوانب المتعلقة بالخمور والأنبذة وطرق تصنيعها ماعدا الأعناب وكيفية نموها وحصادها. والتي تخص فرع أخر وهو الكروم .. منقول يُرجى التدقيق }، ولدينا فن زراعة الدوالي viticultura، وعلم بيولوجيا وزراعة الدوالي ampelología والتي تحولت الى عوالم خاصة قائمة بذاتها. ربما واحدة من الروابط " جينات – نمط ظاهريّ { النمط الظاهري phenotype هي كل ما يصف صفة أو مواصفة مميزة في متعضية، مثل مورفولوجيتها أو شكلها، نموها، أو سلوكها، على عكس النمط الجيني وهي عبارة عن تعليمات مشفرة مورثة يمكن ان يتم التعبير عنها أو تبقى بدون تعبير ظاهري... منقول يُرجى التدقيق } لافضل عنب معروف: هو ما يفسّر اصل تنوع الوانه. كما هو مفترض، فتعرفون بوجود انواع عنب ذات لون اسود، بنفسجي، زهري، ابيض وتدرُّجاتها. ومن المثير أن هذا الامر مضبوط بالكاد من قبل جين واحد اسمه: VvmybA1، عبارة عن منظّم للتوليف الاحيائيّ لبضع الاخضاب المسماة: antocianinas. فلكي يظهر اللون يحتاج قليل من الحيل الجينية. تقوم بضع تغيرات في المنطقة المحفزة لهذا الجين باعطاء المجال لظهور الوان فاتحة وزهرية، الامر الذي يسهل تفسيره، حيث تقوم المناطق المحفزة بضبط تعبير الجين بصيغة أنّ أيّ تغيّر فيها، يمكن ان يدفع الجين لتعبير أقلّ. في حالتنا، يُسبّب تعبير اقلّ بظهور اصباغ اقلّ وبالتالي لون العنب سيكون فاتحاً اكثر. من الذي قال بأنّ كل الطفرات تكون سلبيّة؟ من جهته، يكون اللون الابيض عبارة عن ميزة متنحيّة، ما يجعل تنوعات اللون الابيض هو تماثل الألائل بالنسبة لأليل " ابيض "، وكيف يُولد مثل هذا الجين؟ هل تتذكرون الينقولات التي تحدثنا عن سابقاً؟ " العناصر الوثّابة " ؟ فيظهر عند ادخال " عنصر وثّاب " يسمى Gret1 في المنطقة المحفزة لجين VvmybA1 حالة اعاقة لتعبيره { لقدرته على التعبير }، الامر الذي يقلل من الاصباغ بالنهاية، ما يؤدي لحصولنا على العنب الابيض. وهذا يفسّر سبب كون هذا الأليل متنحي، سيكون كافياً ان يتمكن الأليل من التعبير ليبدأ العنب بمراكمة الاصباغ.  تنوعات بلون العنب واقتضاء حصول تعديلات جينيّة تنوعات بلون العنب واقتضاء حصول تعديلات جينيّةيتبع |
|
|
|
|
|
|
07-31-2016, 07:10 PM
|
رقم الموضوع : [3] |
|
عضو ذهبي
|
البُستان التطوريّ {3}: العالم السوفييتي Karpechenko وانتاجه لجنس نباتيّ جديد، هو: Raphanobrassica  نوع نباتيّ جديد ¡Raphanobrassica! في ربيع العام 1927 وفي معهد علم النبات التطبيقي في مدينة Detskoe Selo السوفييتية، كان العالم Georgii Dmitrievich Karpechenko يتأمّل عمله الاخير. راجع خطواته، دقّق نقطة نقطة، كل شيء كان على ما يرام، لقد انتهى لعمل نوع جديد من النباتات، جنس نباتي جديد هو: Raphanobrassica .. كل شيء بدا كاملاً، لكن لماذا وجب ان تكون النتيجة هكذا؟ وما هو Raphanobrassica ؟ قبل كل شيء هو جنس من النباتات ينتمي لعائلة Brassicaceae والتي تسمى ايضاً الفصيلة الصليبية أو الكرنبية Crucíferas بسبب شكل الزهر الصليبيّ، فيها اكثر من 3700 نوع مُصنّف بحوالي 330 جنس. تكون اغلبيتها عشبيّة ذات دورة حياة سنوية او دورة لعامين، متوفرة ومنتشرة كثيراً في كل مكان تقريباً.  جدول بالميزات الرئيسية لعائلة Brassicaceae =Crucíferas. لاحظوا شكل زهرتها وثمرتها. يكون ملحوظاً أهمية هذه العائلة الزراعيّة، فمن منا لم يأكل أو يسمع بالبروكولي، القرنبيط، الملفوف ... الخ؟ مع كثير من الاسماء والمشاهد المتباعدة، والتي تجعل من الصعب علينا احيانا اعتبارها من ذات النوع، ومع ذلك، هي هكذا فعلاً من ذات النوع. فكل تلك النباتات عبارة عن تنوعات من نوع Brassica oleracea L. بالمناسبة، لا يتوجب الخلط بين الملفوف والخسّ (Lactuca sativa L.)، فبينما ينتمي الملفوف الى الصليبيات، فإنّ الخسّ ينتمي الى المركّبات { فصيلة نباتية من ذوات الفلقتين وحيدة القُعالة سفلية الاسدية وتعتبر من اكبر الفصائل النباتية .. قاموس المورد اسباني عربي }، وهي عائلة من النباتات تضمّ انواع مثل سنّ الاسد، الاقحوان أو البابونج كمثال. يكون نوع صليبيّ شهير ذو اهتمام زراعي كبير هو الفجل. ايضا منتشر بكل مكان ويمتلك تنوعا نسبياً لعدد من الانواع. سبب زراعته هو جذره القابل للاكل والنمو الملحوظ. ينتمي هذا النوع لجنس مختلف عن الجنس الذي ينتمي له الملفوف، هو وفق التسمية العلمية: Raphanus sativus L  نموذج من الفجل نوع Raphanus sativus L حيث تقتصر أهميته للآن في عالم الزراعة على كونه مشروع بحث، واذا ظهر في بستان احدهم فعلى الارجح انه لا يميزه عن عشبة ضارة. ومع هذا، فإنها نبتة بالكاد عمرها قرن من الزمن!! لهذا يواجه تصنيفهاعلمياً شيء من التعقيد، كونها لا تنتمي لنوع ظهر في الطبيعة، بل ظهر في دفيئة مخبريّة زراعيّة. وفي زمن كان علم الوراثة فيه: يمشي خطواته العلميّة الاولى.  مثال من نوع Raphanobrassica. صورة تعود للعام 1929. ولادة ... نموّ بمضي عقد عشرينيات القرن الماضي، يكون قد مضى 30 عاماً على اعادة اكتشاف مستقل لقوانين ماندل الشهيرة من قبل: عالم النبات الهولندي Hugo de Vries، العالم الالماني Carl Correns والعالم النمساوي Erich von Tschermak، أما مُصطلح " جين " فقد وضعه عالم النبات الدانماركي Wilhelm Johannsen الذي لم يبلغ عمره اكثر من 20 عام وقتها، وبمرور 10 سنوات سيقوم عالم الوراثة الاميركي Thomas Hunt Morgan بتحضير ما كان يسمى فرضية " الكروموزومات الحاملة للجينات "، وكان ما كان بذلك " الشيء " الذي أسموه " جين gen " و كان ملمحه الوحيد المعروف: بأنه شكل اللبنة الاساسية للوراثة. هكذا اذاً، شكّلت دراسة الكروموزومات في ذاك الوقت الصرخة الاخيرة في علم حديث اسمه علم الوراثة او علم الجينات بدقّة genética.  العالم Georgii D. Karpechenko (1899-1941). صورة تعود للعام 1929. في ذاك الوقت عمل الشاب Georgii D. Karpechenko كعالم نبات ووراثة في معهد علم النبات التطبيقي في مدينة Detskoe Selo القريبة من لينينغراد. كانت تتركز ابحاثه في عائلة الصليبيات، لقد درس طبيعة كروموزوماتها من الميزات الموروثة وكيف تبرز كلاها في الهجائن. ولم يطل به الوقت إثر انتهائه من دراسة الطبيعة الكروموزوميّة للتهجين، حتى احرز وبنجاح تهجين نوعين Brassica oleracea و Raphanus sativus. الامر الذي لم يكن صعب عليه، فقد اهتم بتلقيح زهرة من نوع Brassica oleracea مع غبار طلع لنوع Raphanus sativus، وبعدها تابع عمله مع تلك الهجائن في المُختبر سامحاً لها بالانتاش والنمو. واجهته عقبات، فكان لديه عدد من الاجنة المُجهضة في الطريق. لكن تمكن عالمنا Karpechenko من تحقيق انتاش ونمو تلك الهجائن. مع هذا لم تكن جديدة كلياً، فقد تمّ تحصيل ما يشبهها سابقاً. فيما لو لم يتم تحقيق مثل هذه الدراسة التي قام بها عالمنا سابقاً، فبرأيي الشخصي ما قام به من تخصيب تهجيني، لم يكن حادثاً عجيباً في الطبيعه. وبفضل شهادات علماء آخرين، فإنّ هجائن عالمنا كانت غير خصيبة. ونحن نعرف بأنّ تهجين نوعين مختلفين { المثال الاشهر الحمار والفرس } حيث يكون المتحدر البغل وهو مُصاب العقم اي غير خصيب. لهذا فإنّ القاح المتحدر الهجين يكون الصيغة الافضل للفصل بين الانواع. في حالة هجين Raphanus xBrassica كحادث يمكن ان يكون اكثر شهرة، لولا انّ التهجين قد جرى بين جنسين مختلفين لا بين نوعين مختلفين { وفق علم التصنيف، يكون مستوى تصنيفها اعلى من مستوى النوع }. ولهذا كان ورود حادث العقم للهجين أكبر من المُعتاد. النباتات مثلها مثل الحيوانات، تحمل في نوى خلاياها مجموعتان من الكروموزومات (2n)، بينما تحمل الامشاج، الخلايا الحاملة لعبء التكاثر { غبار الطلع والبويضات }، مجموعة واحدة من الكروموزومات (n). خلال تشكّل المشيج، يكون مهما ان يتعرّف كل كروموزوم على زوجه الموافق قبل انقسامه النهائي الى خليتين مختلفتين. عملية انقسام اختزاليّ أو تشكيل الأمشاج. تتمثل في النقطتين 1 و2 العملية الجوهرية بالتعارف المتبادل للكروموزومين مع ذلك، تكون الكروموزومات في تلك الهجائن مختلفة فيما بينها، بحيث لا تمكّن من اعطائها هذا الاعتراف. كنتيجة تطورت امشاج ذات عدد غير طبيعي من الكروموزومات وبناء عليه تكون غير قابلة للحياة. رغم ذلك في العام 1927، تمكّن العالم Karpechenko من تجاوز تلك العقبات البيولوجية. لكن كيف؟ من تابع الجزئين الاولين من سلسلة البستان التطوريّ، من المؤكد انه سيعرف الجواب: لدينا ظاهرة تعدد صبغيات جديدة poliploidía وكعاقبة لها، لدينا: نشوء انواع جديدة. في ظاهرة تعدد الصبغيات { لا تنسوا ان الصبغيات هي ذاتها الكروموزومات .. فينيق } تتضاعف كل المادة الوراثية. بصيغة مؤكدة، تشكّل الأم لكل: الطفرات، لظاهرة التغيّرات الجينية الوراثية الاكثر فظاعة والممكن تخيُّلها. تحدث بسبب خلل بتشكيل الامشاج، بصيغة انها بدل ان تظهر بحمولة n كروموزوم، تظهر بحمولة 2n من الكروموزومات. بصيغة انه باتحاد مشيج ثنائي (2n) مع مشيج آخر ثنائي (2n)، تكون النتيجة: لاقحة رباعية (4n)ن والتي لاسباب حياتية، من المعتاد ان تكون في النباتات قابلة كثيراً للحياة.  جدول يبين اصل نوع Raphanobrassica. لاحظوا الفارق بين الثمار المنفتحة والمنغلقة. لاجل ذلك، في الواقع عالمنا لم " يتخطّ " أيّ قاعده بيولوجية. فلقد استفاد من ثغرة في النظام فقط. فكما اشرنا سابقاً، نتجت اخطاء كثيرة في تلك الهجائن خلال فصل الكورموزومات، الأمر الذي شكّل السبب بانعدام خصوبتها. لكن كتناقض ظاهريّ، كان هذا سببا بانتاج امشاج ثنائية بمعدل اعلى من المُعتاد. يشكّل نوع Raphanobrassica حالة خاصة. لكثير من الاسباب. فعلى المستوى الجيني: يكون متعدد المجموعات الصبغية لنموذج غير متعدد. وضمن هذا النموذج، يكون عبارة عن نوع anfidiploide، وهذه الكلمة يوجد سبب لمعناها الحرفيّ " مجموعات كروموزوم -ploidía المُضاعفة (-di-) وبنموذجين مختلفين amphi- "، ما يعني، جينوم متشكّل من اتحاد جينومين مختلفين. وفي حالة هذا النوع الجديد: من الفجل والملفوف. تكون ميزتها الأكثر اهمية هي: قدرتها على التكاثر!! وكيف امكنها ان تكون خصيبة؟ حسناً، يكون الجواب بسيطاً: لأنه عندما تضاعف الجينوم الاصلي للهجين، فقد امتلك كل كروموزوم قريناً قابلاً لتشكيل زوج دون زيادة او نقصان. فلم يكن الناتج لا ملفوف (Brassica) ولا فجل (Raphanus) ولا حتى هجين منهما { لكونه مصاب بالعقم وفق منطق التهجين التقليدي }. إنه شيء آخر، نموذج نباتيّ آخر، لا يكون نوع نباتيّ جديد، بل شيء أكثر، يكون جنس مختلف وجديد! انه ببساطة: Raphanobrassic جدول آخر لاصل Raphanobrassica. جدول تفسيري آخر لاصل Raphanobrassica. لا تكون اوراقه كاوراق الملفوف او الفجل، لكنها تمتلك نسيج كنسيج الملفوف ومنظر شبيه لما لدى الفجل، وتنمو بصيغة نمو الورود الصغيرة. لا تكون جذورها كجذور الملفوف ولا الفجل، على الرغم من انها تذكرنا بجذور الملفوف لكن بالياف اقلّ. كذلك يحصل مع الازهار في الغالب، انها بيضاء ميّالة الى البنفسجي، تذكرنا اكثر بالفجل. وتمتلك الثمرة بالنهاية جزء قاعديّ منفتح مثل الملفوف وجزء طرفيّ منغلق مثل الفجل. بالنهاية، لدينا موزاييك من الخصائص، عدا المرغوب منها. تكون تلك الخصائص من نمط " تظهر لكنها لا تكون ..." كعاقبة لاعادة الترتيب الجيني الناتجة عن تعدّد المجموعات الصبغية الغيريّة alopoliploidía، ظهور جينوم جديد لم يكن موجود سابقاً في الطبيعه كحصيلة اتحاد واعادة ترتيب كامل لجينومين سابقين وبمدى زمني قصير، تكون نتائجه الفورية: اخماد بعض مناطق الجينوم، تنشيط مناطق اخرى وظهور طرق ترتيب جينية جديدة غير موجودة سابقاً. اما على المدى الطويل، فلدينا كمّ هائل من مادة جديدة ستسمح بظهور جينات جديدة. أخيراً، وبكل الاحوال، للآن يتبقى الكثير للعمل بهذا المضمار.  Raphanobrassica، صورة للجنس النباتي الجديد تعود للعام 1929 ومن تصوير عالمنا ذاته  مزايا Raphanobrassica، A الجيل الرابع من النبات، B الجيل العاشر من النبات، C قرنين مع 15 بذرة، D زهور للنبات، E بذور متنوعة له، F وG انقسام اختزالي، H وI تصوير موجة DNA يكون كروموزوم Raphanus sativus باللون الأحمر. يتبع |
|
|
|
|
|
|
08-01-2016, 09:05 AM
|
رقم الموضوع : [4] |
|
عضو ذهبي
|
البُستانُ التطوريُّ {4}: تطوُّر القطن  نماذج لنوع Zea mays subsp. mays L، دقّقوا جيداً بالنبات، ساق وحيد طويل حامل لعرانيس مُمتلئة البحث عن الفروقات الذرة. نبتة متعاظمة اخرى، قد غزت نصف العالم. يستفيد منها كثير من الرئيسيات الحيّة، اسمها في بلدي panizo، اما العلماء فقد سموها: Zea mays ssp. mays L. انتشرت بالعالم جنبا لجنب مع القمح، الشعير والشوفان وعدد من الانواع الاخرة المنضوية تحت اسم عائلة النشويات. تمتلك دورة حياة سنوية ولديها القدرة لبلوغ اطوال حتى 2.5 متر. هي تكون وحيدة الجنس { يحضر الجنسان الذكري والانثوي بذات النبتة } monoica، وهذا يعني انها لا تمتلك ازهار ثنائية الجنس hermafroditas { كما هو معتاد في النباتات }، بل تمتلك ازهار ذكرية من جانب وازهار انثوية من الجانب الآخر.  عرانيس ذرة متنوعة لكن من أين تأتي الذرة؟ بمضيّ اعوام من الدرس وعقود من النقاشات، يتفق اغلب العلماء حول السلف المباشر البري لذرتنا المزروعة في الوقت الراهن. حقّق هذا الامر نبتة محددة مثل Zea mays ssp. Parviglumis { البديل الاكثر قابلية للحياة هو Zea mays ssp mexicana من الاقارب المقربين جداً من النوع السابق }. انها نبتة معروفة على مستوى محلي وعالمي تحت اسم: teosinte، teocinteteosintle أو teocintle. ذات الامر، بكونها نوع فرعيّ بمنتصف الطريق نحو الانقسام لانواع عديدة مختلفة. في الواقع، فيما لو نكن اكثر دقّة، يجب ان ناخذ بحسباننا عدم وجود نوع واحد فقط من teocinte، فهذا المُصطلح قد شاع بالعالم كتعبير عن انواع مختلفة من " الذرة البريّة "، مثل: Zea diploperennis, Zea perennis, Zea luxurians y Zea nicaraguensis والعديد من الانواع الفرعيّة، مثل: Zea mays: ssp. huehuetenangensis, ssp. mexicana y ssp. Parviglumis حيث تشكّل مجموعة صغيرة من النباتات باعضاء ذوي دورة حياة سنوية ودورة سنتين، تجد اصلها في المكسيك واميركا الوسطى. في شجرة هذه الشعبة الوراثية الجينية وفي الشعبة اللاحقة: يمكن تقدير علاقاتها التطورية.  شجرة الذرة الوراثية الجينية وبعض اقاربها بمستوى جينومي كلها تكون ذات الصيغة. انها نباتات ثنائيّة مع 10 ازواج من الكروموزومات فقط في كل جانب. باستثناء نوع واحد هو Zea perennis، فلقد ظهر من تضاعف جيني حصل بجينوم نوع Zea diploperennis ، ما يعني اننا نتحدث عن متعدد مجموعات صبغية آخر { ثلاث او اكثر } poliploide { هذه المرة من نوع رباعيّ 4n }. يمتلك 20 زوج من الكروموزومات اضافة لابرازه عدد من الفروقات نسبة للنوع الامّ بمستوى: الجذمور { ساق ارضية شبيهة بالجذور .. قاموس المورد اسباني عربي }، نمط حياة.  نماذج من teocinte o teosinte ما الذي تمتلكه انواع Zea mays ssp. parviglumis y Zea mays ssp. Mexicana ولا تمتلكه النسخة " البريّة " من نباتات اخرى مزروعة؟ بببساطة النوع المزروع من الذرة والنوع البري: يكونان مختلفان بشكل كليّ، لدرجة امتلاكهما لخصائص متعارضة. فمن جانب لدينا التشريح الذاتيّ للنبات، ففي الذرة: يوجد ساق منتصب طويل مزود بسنيبلات ذكرية ومليء بالاوراق التي يُلاحظ في براعمها تكوّن العرانيس. لكن نرى في الذرة البريّة teocinte ساقاً رئيسياً يتوزّع لتفرعات جانبيّة { وكل فرع جانبي بدوره يحمل سنيبلات ذكرية واوراق مع سنيبلات صغيرة انثوية في براعمها }. فالمتطلبات تكون هائلة هنا، بحيث انه لاجل " تحول " نبات ذرة بري teocinte الى نبات ذرة مزروع، يجب قبل كل شيء : تحويل " فرع جانبي " الى عرنوس.  جدول مقارنة بين الذرة المزروعه والنوع البريّ لكن المشكلة لا تنتهي هنا، فالنوع البري teocinte لا يُنتج عرانيس. يشكّل العرنوس بنية كبيرة، مخروط بطول 10 الى 20 سنتمتر، يحتوي بذور عارية مرتّبة بعدد يبلغ من 200 الى 400 بذرة { ويمكن ان تصل الى 1000 بذرة }. من جانبها، تحتوي سنبلة النوع البري teocinte بالكاد من 6 الى 12 بذرة بحجم صغير، غامقة اللون، بشعة ومحمية بقشرة قاسية جداً. يكون كل شيء بمثابة النقيض.  صورة مُقارنة بين النوع البريّ سنبلةفي الاعلى ولونها اخضر،) والذرة (عرنوسبالاسفل ولون اسود ليس بمُستغرب اعتبار تطور النوع البري teocinte كواحدة من الظواهر الاكثر تطرفا وتعقيداً للتطور المورفولوجي الممكن العثور عليه بالوقت الراهن. فلقد فكّر بعض الباحثين بأن النوع البري للذرة teocinte: قد كان اكثر قرباً من الرز منه من الذرة، مع ذلك فهجائن الذرة تكون بالعموم ناجحة وخصيبة بشكل كامل. وضعت الدراسات الجزيئية الحديثة النقطة الاخيرة في النقاش حول هذا الموضوع.  جدول مقارنة آخر بين النوعين البريّ والمزروع: A فروقات في التفرعات الجانبية للنوع البريّ الحامل للسنيبلات الذكرية، B مقارنة بين السنيبلة والعرنوس، C مقارنة بين السنيبلات الصغيرة والاغطية الحامية قضيّة تشريح هنا يحضرنا التساؤل: كيف ظهرت الذرة المزروعة اعتباراً من الذرة البريّة teocinte؟ لا تكون قضيّة سهلة الحلّ. تابعت اوائل الاجوبة " منطق الفروقات الضخمة، ثمّ كميّة الجينات الضخمة اللازمة ". هل حصل تطفُّر لعدد غير محدد من الجينات في العشر آلاف عام الاخيرة { يمكن افتراضه كتاريخ لبدء تدجين الذرة }؟ أيكون اصل الذرة اقدم بكثير مما نتخيّل؟ لقد اثبتت ابحاث الاربعين عام الاخيرة والتقدّم الحاصل بعلم الاحياء الجزيئي: بأنّ الفروقات الرئيسية بين الذرة الزراعية والذرة البريّة teocinte، تعود الى وجود خمس مجموعات من الجينات، او من الافضل القول خمس QTL (quantitative trait loci). حيث يشكّل QTL: التسمية التي تستقبلها تلك المجموعات من الجينات التي ترتبط بقوّة لاجل التوريث معاً، والتي تساهم ايضا ببناء ذات الميزة. بفضل اعمال عالم الوراثة الاميركي John Doebley نعرف النوع (teosinte branched1 (tb1، تكون وظيفة QTL منع النمو، اعاقة نمو التفرعات الجانبية { مع اوراقها، سنبلاتها الذكرية وسنبلاتها الانثوية }. كيف يتصرف tb1 ؟ حسناً، لنقارن فعله في النوع البري teocinte وفي الذرة الزراعية: أولاً: في البراعم الجانبية للنوع البريّ teocinte، يكون تعبير tb1 قليلاً أو مُنعدماً، بالتالي لا يظهر شيء بمستوى جزيئي يقوم بمنع نمو التفرعات الجانبية، وبالتالي تنمو تلك الفروع. ثانياً: مع هذا يعبّر هذا الجين في الذرة الزراعية بسرعة مُضاعفة لسرعته في النوع البري، كنتيجة لرسالة " منع النمو " تنتشر في البراعم الجانبية، وكنتيجة لا تنمو التفرعات. لهذا السبب، تقوم اغلب نباتات الذرة المتطفرة ب tb1 عند فقدان وظيفية QTL بتوليد افرع جانبية ذات حجم أكبر. بالنهاية من المعروف بأنّ النشاط المُختلف ل tb1 في النوع البري والنوع الزراعي، لا يعود لكون كلا الجينان مختلفان. كلا ففي الواقع يكون الجين tb1 في النوع البري والنوع الزراعي هو ذاته. ما يحدث في كلا النباتين، هو أن ترتيب الجين يكون مُختلفاً، الامر الذي يقود الى ايقاع نشاط مختلف لكل نبات منهما. يقتضي اختلاف ايقاع التعبير الجيني أنّه في النوع البري، يكون هذا الجين اقلّ نشاط بوظائفه، وعلى العكس من هذا في الذرة المزروعه، حيث يكون الجين أكثر نشاطاً بكثير. الامر الذي يثبت لنا بأنّه لا يهم حجم او دقّة التعديل في جين واحد فقط باتجاه دقيق لاجل احراز فروقات مورفولوجية ضخمة.  A نبات ذرة بريّ teocinte Zea mays mexicana C النوع المزروع Zea mays mays E عيّنة الذرة المزروعة المتطفرة tb1 بما يتصل بما سلف، تمّ العثور على QTL آخر يسمى: barren stalk1 ba1، كما لو كان مرآة تبيّن لنا ثقتنا بانفسنا، كما لو كان الصيغة التي تحوّل الشاكي Jekyll الى الهمجيّ Hyde، يكون النوع barren stalk1 المُعاكس الخفيّ للنوع البري teosinte branched1. ففيما لو رأينا الجين tb1 المتعرض لانخفاض نشاطه: حيث نرى نباتات بتفرعات جانبية هامة، فإننا سنجد بأنّ تخفيض نشاط الجين ba1، سيؤدي الى نباتات دون تفرعات جانبية { على الرغم بأنه في الواقع، تمّ العثور على طفرات بدرجة واسعة في هذا النوع من التعديلات } بنوع من الثقة نقول بأنّ الجين ba1: يشكّل منظّم ضروري في التشكيل المبكر للبراعم التي تعطي السيقان الجديدة { سيقان جمع ساق }، الازهار او الثمار الجانبية. لهذا تُرى طفرات هذا الجين مصغّرة لنمو التفرعات الجانبية من جانب, مبيّنة من الجانب الآخر نمو عمودي قويّ للنبات. تشكّل تلك الثنائية: قاعدة الشُبهة في التوازن بين ba1 و tb1، وتكون السبب المُحقق لراحة البنية التشريحية للذرة الحديثة.  مؤثرات طفرة الجين ba1على نمو الذرة.a : ازهرار ذكري بنبتة متطفرة، b : ازهرار ذكري بنبتة برية، c وd:فروقات في النموّ التكاثريّ والإنباتيّ،على التوالي،بين نباتات برية الى اليسارومتطفّرة الى اليمين، لاحظوا الافتقار الى البراعم الجانبية في الاخيرة يوجد فارق بارز آخر، تكون بذور النوع البري teocinte محمية بقشرة سميكة، غامقة ومتخشبّة، وهي حماية غائبة في الذرة المزروعة ومُختصرة الى مقعد ضخم للبذور العارية. تُشير دراسات جزيئية عديدة لأنّ واحد من QTL الاكثر أهميّة لنمو تلك الاغطية هو جين teosinte glume architecture1 tga1، والذي تحقّقت تقدمات اكثر اهمية به من قبل فريق البحث Wang, H. et al في العام 2005. فنتائج ابحاثه المنشورة بمجلة الطبيعة: تستخلص بأنّ tga1 عبارة عن جين مُنظّم آخر ويعمل كنقطة انطلاق في تشكيل تلك الكؤيسات. ايضاً لاحظوا وجود 6 فروقات جينية بين جين الذرة الزراعية tga1 وبين جين الذرة البرية tga1، والتي من بينها: تغيّر في المنطقة السادسة بسبب النشاط المُختلف للجين في نبات أو آخر. بمطابقة نموذج الباحثين، نلاحظ بكفاية طفرة واحدة في جين اساسي لاجل اعطاء الاصل لبذور عارية نراها في الذرة الزراعية انطلاقا من بذور محمية بقوّة للنوع البري teocinte.  a : عرنوس ذرة يظهر محوره، b: عرنوس بري teocinte، c: ذات العرنوس البري مع أليل tga1، d: بذرة نوع بري، لاحظ نموّ القشرة الخارجية، e: بذرة للنوع البري مع الأليل tga1، f: عرنوس ذرة مزروع، لاحظ القشور، g و h: عرنوس بري مع الأليل tga1 استطالت القشور وتغلفت بعض البذور أصول العرنوس يتبقّى علينا شرح الملامح الكبرى المتصلة بالبنية التشريحية للذرة الزراعية وبذورها العارية، ما يعني القيام بشرح اصل العرنوس ذاته، نموه وانتشاره، بالقياس مع سنبلة النوع البري teocinte. لا يكون حل هذه القضيّة خالي من المشاكل، فللآن لا يوجد معطيات قطعية ولا خلاصات نهائية، على الرغم من توفر معلومة كافية قادرة على اعطائنا فكرة عمّا حصل. يعود الدليل الاقدم على الذرة الزراعية الى كهف Guilá Naquitz في المكسيك. عبارة عن مكان مليء بالادغال الكثيفة والواقعة على ارتفاع 1926 متر عن سطح البحر، مع هطولات مطرية لا تتعدى 600 مليمتر. اثبتت الحفريات حضور نشاطات بشرية لصيادين وملتقطي ثمار، منذ الفترة الواقعة بين 10650 الى 6980 عام قبل الآن حسب تقنية تأريخ الكاربون 14.  صورة تُظهر لنا مكان وادي Tehuacán وكهف Guilá Naquitz ، مكان التوزّع الحديث لسلف الذرة Z. mays parviglumis تمتلك باقي انواع الذرة الاكثر قدماً في طبقات كهف Guilá Naquitz عمراً { محسوب عبر قياسات دقيقة بالكاربون 14 } لفترة تعود الى 5.420 ± 60 و 5.410 ± 4040 عام. ايضا عثروا على حبيبات من غبار الطلع لنوع Zea بعمر 9.500-6.980 عام، وكأدلة على زراعة اليقطين Curcubita pepo لفترة تعود الى 8.990-7.000 عام قبل الآن. لم يعثروا بين تلك البقايا على العرانيس التي نعرفها، لكنهم شاهدوا " ميني عرانيس " بميزات شديدة الخصوصية ولا يبلغ طولها أكثر من 25 ملمتر وهو حجم مشابه لسنابل النوع البري teocinte Z. mays ssp. Parviglumis.  تصوير لعرانيس الذرة المعروفة الاكثر قدماً والمُكتشفة في كهف Guilá Naquitz. في المكسيك، العرنوس الاكبر في الاسفل طوله 25 mm يشكّل العرنوس بُنية خصوصيّة. فهو يمتلك محوراً صلباً ومرناً تنتظم عليه الحبيبات العارية بصيغة صفوف " مضاعفة ". بدورها الحبيبات تتموضع عمودياً على محور العرنوس الطولاني. من جانبها، تمتلك سنبلة النوع البري teocinte محوراً تنتظم عليه الحبيبات المحمية بقشرة قاسية بصفين " بسيطين " فقط بوضعية موازية للمحور الطولانيّ. بدورها لا تنتمي عرانيس كهف Guilá Naquitz لكلا النوعين، بل تمتلك ميزة خاصة انتقاليّة. انها متكونة من محور تنتظم عليه بذور عارية، بطريقتين: الاولى على طول صفين " مضاعفين " ومتشكلة بزاوية 45 درجة نسبة للمحور الطولاني { وليست موازية كما في النوع البري teocinte أو عمودية كما في الذرة الزراعية }، والثانية متشكلة من صفين " بسيطين " ومتوجهة بزاوية 180 درجة نسبة للمحور الطولانيّ. هذا المزيج من الميزات { بعضها خاص بالنوع الزراعي، وبعضها بالنوع البري وبعضها الآخر متوسط } يوضح لنا حقبة كانت فيها ميزات الذرة الحديثة في بداية الطريق التكوينيّ.  تنوّع حاضر في عرانيس الذرة الاقدم المعروفة، المُكتشفة في كهفGuilá Naquitz فيما لو تكن لُقى كهف Guilá Naquitz المدخل للموضوع، فإنّ لُقى وادي Tehuacán في المكسيك ايضاً: تشكّل العقدة وحلّها.فنجد في تلك المنطقة آثار بشرية وبقايا لذرة بدائيّة. فكما في كهف Guilá Naquitz، نلاحظ في الطبقات الاكثر قدماً عرانيس بطول يتراوح بين 19 و25 ميلمتر مع 4 الى 8 حبيبات، مع ذلك، ترتفع الطبقات في تلك المنطقة حتى تصل لوقتنا الراهن، الى ما قبل 500 عام من الآن. خلال كل تلك الحقبة، نواجه التعاقب التطوريّ الكامل لعرنوس الذرة مع الاسلاف من العرانيس الصغيرة في الطبقات الاكثر قدماً والعرانيس الحالية في الطبقات الحديثة.  التعاقب الكامل لتطور عرنوس الذرة منذ 7000 عام قبل الآن وصولاً الى 500 عام قبل الآن وفق ما سلف، يمكن التأكيد على أنّه في ظهور وتطور العرنوس، تكون لحظة الصفر هي النقطة الاكثر عرضة للنقاش. ونعود لبداية كل شيء، ما الذي حوّل تشعباً { تفرّعاً } للنوع البري teocinte الى عُرينس صغير اصليّ ؟ بحسب اعادة بناء بقايا احفورية، فإنّ العرينس الاصلي: ظهر عبر بنية لا تكون اكبر بكثير من سنبلة النوع البري teocinte.  مقارنة بين A سنبلة النوع البري teocinte ، B نوع الذرة البدائي الاكثر قدما،C نوع الذرة الحديث تمّ اقتراح عدد من التفسيرات لحلّ هذه الاحجية. يتطرق بعضها لنوع من التهجين، حيث سيكون نوع الذرة البدائي ثمرة عملية تهجين بين النوع البري teocinte من جنس Zea ونوع آخر من الجنس Tripsacum { وبينهما علاقة قرابة لصيقة جداً }. يدافع اصحاب هذا الاقتراح على وجه الخصوص عبر الشبه الهائل لعرنوس بعض الهجائن بين Tripsacum و teocinte، والذي من المُفترض انه شكّل العرنوس البدائي للذرة، نقطة اخرى بسبب حضور تعدّد الاشكال الخاصة بالنوع البري teocinte وبنوع teocinte في الذرة، مع ذلك، لا يخلو هذا النموذج من المشاكل، كعدد كروموزمات الهجائن { المختلف عن 10 } أو ضعف الخصوبة وضعف النجاح بالتهجين بالتالي.  هجين F2بين النوع البري teocinte و نوعTripsacum ، لاحظوا الشبه الهائل مع النوع البدائي للذرة في الرسم. اقتراح بديل للسابق، قدّمه عالم النبات الاميركي Hugh H. Iltis عبر " النظرية الكارثيّة للتحول الجنسيّ " Teoría Catastrófica de la Transmutación Sexual CSTT، وبحسبها: يظهر عرنوس الذرة بسبب تبدُّل في الفيزيولوجية الهرمونية، والتي تنتهي بافاضة الهرمونات الانثوية على السنابل الذكرية، محولة اياها الى عرانيس. في الوقت الراهن، لا يوجد اي اهتمام بهذا الاقتراح. مع ذلك، يقترح عالم النبات ذاته تحديث لنظريته لتصبح " النظرية الجديدة للتحول الجنسي " Nueva Teoría de la Translocación Sexual STLT، وفق هذا النموذج: فيما لو تقترب السنابل الذكرية بوفرة من الازهار الانثوية، فان الهرمونات الانثوية ستنتهي بقلب جنسانية السنبلة الذكرية { اي تحويلها }. هذا بدوره سيجد اساسه في الانتقاء المتقدم للنماذج مع عرانيس كبيرة، يتأسس حجمه في اقترابه الاكبر من ساق النبات. جدول موجز ومعدللنموذج STLT لعالم النبات Hugh H. Iltis لاجل تفسير اصل عرنوس الذرة، 1 فرع جانبي مع سنبلات انثوية محورية وسنابل ذكرية طرفية 2 تحول جزئي لسنبلة ذكرية طرفية الى سنبلة انثوية، 3 انتقاء السنبلة الانثوية الطرفية، 4 سيطرة للسنبلة الانثوية الطرفية. ربما يكون هذا النموذج معقولاً، وهنا ندخل في نموذج ثالث للخلاف، " الفرضية المُجمع عليها للنوع البري Teocinte " (OTH)، وبحسب هذا النموذج فإنه بفضل طفرات اساسية بجينات اساسية، أُعطي الاصل ليس فقط للجسم التشريحي للذرة، بل ايضاً لتكوين عرانيسها. فيما لو نتذكر الطفرة tb1 فإنّ الذرة المتطفرة قد طورت سيقان طويلة جانبية شبيهة بما لدى النوع البري teocinte، لكن ليس هذا فقط، اضافة لانه في اطراف تلك الفروع الجانبية نمت سنابل ذكرية كما في النوع البري teocinte { وليست انثوية كما في الذرة }. مع هذا، يمكن الذهاب ابعد من هذا، عندما يصطدم مع اقتراح Iltis. جرت ملاحظة أنه عند حدوث " طفرات مضاعفة " واحدة منها tb1 والاخرى dwarf1 d1 أو anther ear1 an1 طفرات تساهم بانتاج نباتات قزمة، في الفروع الجانبية تتابع السنبلة الذكرية تشكّلها. هذا يُشير لأنّ تشكيل السنبلة الذكرية لا يتوقف على طول الفرع الجانبي، بل هو عبارة عن ميزة مستقلة. فكما رأينا، على الاقل بجزء " تطور معكوس " من الذرة الى النوع البري teocinte، هل يوجد لُقى اكثر بفضل النموذج OTH؟ باتجاه واضح، نعم. دون الذهاب بعيداً، نعرف بعض الطفرات, مثل ts2 وهي قادرة على التسبّب بانهاء نمو الازهار الذكرية الى ازهار انثوية. ليس هذا فقط، ايضا أكّدت بأنّ الطفرات المضاعفة tb1: ts2 تحقق السنابل الذكرية جزئياً أو السنابل الانثوية كليّاً.  في الصورة (A) تفرعات جانبية للطفرة المضاعفة tb1 : ts2 للطفرة tb1 والطفرة المضاعفة tb1 : an1 يمكن ملاحظتها كطفرة مضاعفة فقط tb1 : ts2 تبيّن تأنيث السنبلة الذكرية، في الصورة (B) نمو اكبر في شعيرات النبتة المتعرضة للطفرة المضاعفة ts2 : tb1 بل اكثر من ذلك، في وقتنا الراهن، يجري اختبار حول امكانية تحوله الى نوع Zea mays ssp. Parviglumis بمدة قصيرة، عبر ضغط انتقائيّ في نموذج نباتي يذكّرنا بنوع الذرة البدائيّ. تبين نتائج الاختبار المعمول في جامعة Wisconsin بأنه منذ الحلقة الاولى يمكن رؤية: نباتات بتفرّع طبيعي، نباتات بتفرّع غير موجود تقريباً ونباتات بتفرّعات متوسطة. ليس هذا فقط، بل ظهرت سنابل ذكرية مؤنثة جزئياًن والتي تُذكِّرنا بصيغة مؤكدة " بالذرة البدائية " والتي شاهدنا بقاياها " الاحفورية " سابقاً.  سلسلة من صور الاختبار المسمى Doebley Lab حيث نلاحظ التناقص التدريجي للتفرّع الجانبي بالنوع البري teocinte كما في الصور a, b و d c يمكننا رؤية سنبلة ذكرية مؤنثة جزئياً بالاضافة لملاحظة كيفية تغيّر النوع البري teocinte بيئياً بفضل ميزات الوسط الموجود فيه، فهو يُكيّف تشكّلات مختلفة. الكثير منها يذكرنا بالذرة الحديثة:  جدول يبين التكيفات البيئية للنوع البري، A مثال من نوع Z. m. parviglumis التي تنمو في منطقة Balsas في الشروط المفضلة للنبتة، B نموذج من المنطقة العليا في Missouri شبيهة بنماذج وادي Río Grande في المكسيكتكون اكثر ارتفاع واقلّ تفرعات،فيها عرانيس باربع حبّات، C نماذج شمالية مع عرانيس بثماني حبيبات، من المفترض انها انتشرت من تكساس والمكسيك، Dذرة حديثة. التدجين تمّ التقدّم كثيراً بمحاولات فهم " كيف " تطورت الذرة من النوع البري teocinte. لكن، ما دور البشر في تأصيله؟ اسباب تدجينه؟  أحد آلهة الذرة Yuum Kaax ويعني اسمه " سيّد الذرة اليافع "! وفق الدليل الآركيولوجي، يُشتبه بحدوث تدجين الذرة في الفترة الممتدة بين 5000 الى 10000 عام قبل الآن. لا يكون غير معقول التأكيد على انّ بدايات تدجينها لتغزو العالم منذ 9000 عام، قد كانت في وادي Río Balsas في جنوب المكسيك. دون ذهابنا بعيداً، فمصطلح teocinte يأتي من كلمة Nahuátl والمستخدمة من قبل شعوب اميركا القديمة للتعبير عن " حبّة الآلهة "، ويقصدون بها النباتات السنوية، مثل: Zea luxurians y Z. mays o las perennes Z. perennis y Z. diploperennis تعود اللقى الاحفورية الاكثر قدماً لطبقات وادي Tehuacán وكهف Guilá Naquitz والتي تعود للفترة الممتدة بين 6000 الى 5000 عام قبل الآن، مع حضور عرانيس مختلفة عن العرانيس الحالية وبميزات مشتركة للذرة الحالية والنوع البري teocinte، وجرت ملاحظة هذه الميزات في اكتشافات حديثة في وادي Río Balsas، والتي تشير لحضور الذرة الزراعية المدجنة منذ 8700 عام على الاقل. يتبع |
|
|
|
|
|
|
08-06-2016, 10:39 AM
|
رقم الموضوع : [5] |
|
عضو ذهبي
|
البُستانُ التطوريُّ {5}: تطوُّر القطن الجزء السابق: تطور الذرة لا القطن!   لماذا يكون القطن ذو فائدة جمّة، ليس لقسم من البشر بل لكل العالم. لا يكون مُختلفاً عن القوميات، العقائد أو الثقافات. ففي الوقت الراهن يُستعمل بكل نماذج الثياب، يُستخرج من بذوره زيت هام ذو فائدة في صناعة مستحضرات التجميل، عدا كونه مصدر للسيللوز المستعمل بتصنيع كافة انواع النقود الورقية، دولارات ويورو. لكن القطن ايضاً هو ابن علم الاحياء البيولوجيا، ويشكّل ثمرة الظاهرة الكافرة المسماة بالتطور!! نحصل على كنزنا الثلجيّ من شجيرة القطن، وهو عبارة عن نبات من جنس Gossypium. كانت الانواع الاكثر زراعة تقليدياً في اوروبا من هذا الجنس، مثل: Gossypium arboreum L موطنه الاصلي جنوب آسيا Gossypium herbaceum L أتى من جنوب افريقيا والتي تقهقرت زراعتها بوقتنا الراهن بسبب نوعين مسيطرين على السوق حالياً، هما: Gossypium hirsutum L اصله من اميركا الوسطى، منطقة الكاريبي وفلوريدا Gossypium barbadense L مصدره اميركا الجنوبية واللذان تحولا لنوعين مفضلين عند المُزارعين. زهرة شجيرة القطن نوع Gossypium hirsutum L يكون في الواقع جنس Gossypium اكثر اتساعاً مما هو مُتوقّع. فهو يضمّ 50 نوع من النباتات، يوجد بينها صيغ شجريّة، جنيبيّة وعشبيّة، حيث جرى استثماره مرات كثيرة في التاريخ البشريّ. ينتمي هذا النوع النباتيّ الى عائلة Malvaceae التي تتضمّن بين نباتات اخرى: الخبّازة malvas Malva sp. y Lavatera sp و النبات الشهير hibiscos Hibiscus sp المنتشر في كثير من الحدائق نظرا لجمال ازهاره. الى اليسار ازهار لنوع Lavatera cretica ، الى اليمين ازهار لنوع Hibiscus sp ينتمي كلا النباتين لعائلة Malvaceae وهي ذات عائلة القطن algodón ابن المسافرين أيّة دروس في علم الاحياء التطوري تُعطينا شجيرة القطن؟ كما يقول بعض محبي استعمال علم المصطلحات " الخاصة "، نعثر في شجيرة القطن على ظواهر " ميكرو تطور microevolución " كما على ظواهر " ماكرو تطور macroevolución "، لهذا فيما لو نرغب بفهم التطور بمستوى نوعيّ، اضافة لظواهر نشوء الانواع المذكورة سابقاً، سيكون بالامكان تحقيق الدراسة بفضل حوادث التطور الجزيئيّ. تكون شجيرات القطن نباتات ثنائية المجموعة الصبغية diploides (2n)، اي تتكوّن خلاياها فقط من مجموعتي كروموزومات { مثلنا }. تبعا لميزاتها التشريحية ولميزات جينومها الخاص، تمّ تضنيفها في 8 نماذج كبيرة، هي: A, B, C, D, E, F, G و Kحيث تمتلك كلها 13 زوج من الكروموزومات في جينومها اي (2n = 26) . يكون توزيع هذا الجنس عالميّ. نعثر في استراليا على مجموعات C, G و K، ونعثر في افريقيا وآسيا على مجموعات A, B, E و F، ونعثر في المكسيك, البيرو وجزر غالاباغوس واريزونا على المجموعه D.  التنوع الهائل للقطن، A و B بذور ناضجة لانواع G. hirsutum مزروع وبري على التوالي، C نوع G. tormentosum من هاواي، D نوعG. arboreum مزروع ينتمي الى المجموعه A ، E نوعG. herbaceumبري عضو في المجموعه A ، F نوع G. raimondii، G نوع G. davidsonii، H نوع G. longicalyx، I نوع G. anomalum ، J نوع G. sturtianum، K صورة يبدو فيها مقارنة لكل هذه الأنواع نصل لما يكون ممتعاً. لو نقم بتذكُّر أن الانواع التي وصلت الى السوق العالمية، هي: G. hirsutum ، G. barbadense فإلى أيّ جماعة ينتميان؟ لا ينتميان الى ايّ من الجماعات المُشار لها سابقاً. كلاهما ينتميان الى مجموعة خاصة، مجموعة grupo AD المتوطّنة في القارة الاميركية، وفيها نجد 5 انواع. ما هي المجموعة grupo AD؟ منذ زمن غير قليل، وفي هذه السلسلة من المواضيع، عرفنا نبات ذو جدول اسماء جينيّ شبيه، انه قمحنا Triticum aestivum L، كما عرفنا عملية تشكّل قوة تطورية فعّالة في المملكة النباتية، انها تعدّد { أو تغاير } المجموعات الصبغية الغيريّة alopoliploidía . فكما راينا سابقاً، تكون عملية تعدد المجموعات الصبغية الغيرية alopoliploidía عبارة عن ظاهرة، يحصل فيها تهجين في نوعين مختلفين، فتعطي نوع هجين مصاب بالعقم، حين انّ لاخصوبته تقود لعدم توافق المادة الوراثية لكلا النوعين، الامر الذي يمنع تشكيل البويضات وحبيبات الطلع القابلة للحياة. مع هذا، احيانا يتسنى لبعض تلك الهجائن انتاج بويضات وحبيبات طلع مع حمل مُضاعف من المادة الوراثية. في حال صادف هذا الوجود واحد من حبيبات الطلع يلقح واحدة من تلك البويضات، هنا يمكن للالقاح ان ياخذ دوره حاملاً لنوع جديد من الكائنات، لا يكون مثل ايّ من النوعين السابقين، ولا حتى مثل الهجين من كليهما، بل هو نوع جديد بميزة alopoliploide قادر على اعطاء نسل خصيب، ويكون عادة غير قادر على التكاثر مع انواع اقارب. ظاهرة alopoliploidía ، A نوع آ، B نوع ب، C نوع هجين، D انتاج هجين وتكوين أمشاج diploides، E القاح امشاج diploides وولادة تغاير المجموعة الصبغية Alopoliploide في الواقع، يعني مُصطلح alopoliploide مجموعة الكروموزومات (-ploidía) المتعددة (-poli-) والغيريّة (alo-)، ما يعني، جينوم متشكّل من عدد من الجينومات المختلفة حسب تضاعف اصل هذا الجينوم. مع ذلك، لا يكون تهجين محض. في عملية تشكيل لمتعدد مجموعات صبغي غيري alopoliploide، سيُعاني الجينوم من تغيّرات هامة، بحيث يمكن اعتباره جينوماً جديداً، لم يكن موجود سابقاً في الطبيعه. بمدى زمني قصير، تخمد بضع جينات وتعبّر جينات اخرى، يتعدّل ترتيب العديد من الطرق الجينية، حتى يمكن ان تصل لتعيد ترتيب مناطق كاملة من الجينوم. بمدى زمني طويل، تكتسب الجينات المختلفة وطرق ترتيبها وظائف اكثر تخصصاً، آليات تعبير جيني نوعيّة اكثر أو حتى الوصول لصياغة وظائف جديدة كليّاً. اذاً تمتلك المجموعة grupo AD للجنس Gossypium اصلاً في واحدة من تلك الظواهر. ينتمي احد الجينومات للمجموعة grupo A { ذات المجموعه التي ينتمي لها الانواع المزروعة تاريخياً في Eurasia، وهي - G. arboreumG. herbaceum }، المتوطنة في القطاع الافريقي الآسيوي، بينما ينتمي الجينوم الآخر للمجموعة grupo D المتوطنة في القارة الاميركية. ما يعني ان المحيط يفصل كلا المجموعتين! تطور وتدجين القطن algodón تُشير الاحرف للمجموعه الجينية التي تنتمي لها كل صورة. لكن يجب الاعتماد على الادلة. يتكوّن جينوم المجموعه grupo AD من 26 زوج من الكروموزومات بخلاف 13 زوج من الكروموزومات التي نجدها بباقي الجنس Gossypium. منذ العام 1940، دراسات التشكيل الجينيّ والتزاوج الصبغي، دراسات كيمياء النبات، تحليل الخرائط الوراثية والتقنيات الاحدث في السلسلة: لم تقم بعمل شيء أكثر من التوافق مع ميزة تعدد المجموعات الصبغية الغيرية alopoliploide لهذه المجموعة، يكون الجينوم AD متشكلاً من جينوم A وجينوم D. ليس هذا فقط. ففي العام 1940، جرى تبيان اصل تلك المجموعه مخبرياً. حيث قاموا بتهجين اعضاء آسيويين من المجموعه grupo A مع اعضاء اميركيين من المجموعه grupo D ولاحقاً ضاعفوا جينوم الهجائن الناتجة، فكانت النتيجة تكوين " متعدد مجموعات صبغية غيرية alopoliploides نوع AD "، اظهرت تلك alopoliploides ميزتين هامتين، هما: أولاً: امكنها التهجين فيما بينها ذاتها واعطاء الاصل لبذور قابلة للحياة ثانياً: امكنها التهجين مع الانواع الطبيعية للمجموعه grupo AD وافساح المجال لظهور متحدرين خصيبين! هذا لا يشكّل دليل على ميزة تعدد المجموعات الصبغية الغيرية alopoliploide للمجموعه grupo AD فقط، بل يشكّل تصنيف لانواع لم تكن موجودة في الطبيعه سابقاً .. يكون مؤسس من جديد في المُختبر. رغم هذا، يكون مرجحاً للآن بقاء بعض الشكّ. وسيكون هذا الامر جدُّ طبيعي، حيث اننا نتكلّم عن كل صنف ضمن الجنس Gossypium، الذي يجد اصله في تهجين صنف افريقي آسيوي مع صنف آخر امريكي. امام امكانية كتلك، تبقى قضيّة لا مفرّ منها، من اين كل هذه القدرة على الانتشار لشجيرات القطن؟ فكما يظهر، كثير أو أكثر. نجد مثلاً سلائل انتشرت من موطنها المكسيك وصولا الى البيرو كنوع Gossypium raimondii، ومن شمال المكسيك الى جزر غالاباغوس نوع G. klotzschianum، ومن غرب الجنوب الاميركي الى جزر غالاباغوس نوع G. darwinii. لقد بلغت بعض اعضاء هذا الجنس جزر هاواي حتى، تائهة بنصف المحيط الهادي ومتحولة الى نوع G. tormentosum بوصفه نوع مستوطن. بالتالي نحن بمواجهة جماعة من المُسافرين. يمكننا قراءة ابحاث علمية مختلفة تدور حول امكان بقاء العديد من انواع الجنس Gossypium طافية لاشهر عدّة بمياه مالحة، مُحافظة على قدرتها الانتاشيّة، بينما يُبدي اعضاء المجموعه grupo A { أب المجموعه grupo AD } احتمالا كافيا للملوحة والطفو كما لاجل البقاء على قيد الحياة بسفر يقطع المحيط. لكن دون ادنى شكّ، النوع الاكثر احتمال، لتلك الظروف، من الشجيرات هو شجيرات هاواي من نوع G. tormentosum والتي تتحمّل البقاء 19 شهراً في المياه المالحة وتبقى قادرة على الانتاش. يرى بعض الباحثين ان المدة يمكن ان تصل الى 3 سنوات. بكل اسف، حاليا يكون بعض تلك الانواع مهدّد بالانقراض، فالخطر الاكبر الذي يواجه النوع G. tormentosum هو التلوث الجيني الآتي من ادخال نوع archiconocida G. hirsutum الى هاواي، يُعاني من ذات المشكلة نوع G. darwinii في جزر غالاباغوس. الانواع الشقيقة Gossypium klotzschianum و Gossypium davidsoni تصل جنوب كاليفورنيا مع جزر غالاباغوس. يتم اقتراح الانعزال الجغرافي كعامل محرّك لنشوء انواع جنس Gossypium. فقد طرح بعض الباحثين من زمن بعيد بأنّ اصل متعددات المجموعات الصبغية الغيرية alopoliploides، يعود الى زمن يسبق انفصال افريقيا عن اميركا الجنوبية، وهذا يُبعد مشكلة كِبَرْ المسافة البحرية الحالية. فيما لو يثبت هذا الامر، فاننا سنكون امام الاصل المُفترض للصنف AD في المدى الزمني القائم بين العصر الكريتاسي وصولاً لاوائل العصر التيراسي { 100 الى 60 مليون عام }. فيما يرى باحثون آخرون أن البشر ونشاطاتهم عبر الهجرة هي من تحمّلت نقل وايصال انواع شجيرات القطن مجموعه grupo A الى القارة الاميركية، وهذا يعني اننا نعود لبدايات الزراعة في العالم { منذ 6000 عام على الاكثر }. في الوقت الراهن، نمتلك تقدّماً في علم الاحياء الجزيئيّ، وقالت ابحاث متصلة بهذا كلمتها بخصوص اصل متعددات المجموعات الصبغية الغيرية alopoliploides والذي يعود لاواسط عصر Pleistoceno { بين 1 الى 2 مليون عام }، عندما انطلق اوائل الجنس البشري Homo من افريقيا. بينما تؤكد دراسات اخرى بأن المجموعه grupo AD لجنس Gossypium، تشكّل مجموعه وحيدة السلك monofilético، بكلمات اخرى، تمتلك كل انواع المجموعه grupo AD اصل وحيد في متعدد مجموعات صبغية غيرية سلف alopoliploide. ثمار القطن تنتظر القطاف تبعات ظاهرة تعدّد المجموعات الصبغيّة poliploidía لم يتوقّف العلماء عند ذاك الحدّ. بمجرّد اكتساب كل انواع شجيرات القطن لظاهرة تعدد المجموعات الصبغية الغيرية alopoliploidía، رغبوا بمعرفة الأثر الذي تفترضه تلك الظاهرة على صعيد تطور تلك النباتات. عندما يتضاعف جينوم ما، يكون متاملاً العثور على جينات مُضاعفة تُحافظ على استقلالية تطورية مؤكدة، وانّ بعضها يمكن أن يتحول إلى جينات زائفه غير قابلة للحياة، جينات أخرى تُحقق وظائف جديدة وحتى القفز من جينوم لآخر. هكذا نجد دراسات على مدى أكثر من 10 أعوام، جرى تحقيقها على 16 نوع Gossypium hirsutum { شجيرة القطن الاكثر زراعة والممثلة للصنف AD }، وقد بيّنت تلك الدراسات بأنه ومنذ ظهور هذا الصنف، بالكاد قد ظهرت جينات زائفة، الامر الذي لم يؤدي لحدوث خسارات جينية وظيفيّة هامة. تؤكد دراسات حديثة محقّقة على 40 جين ومضاعفاتها، عبر مقارنة لشجيرات قطن AD بريّة وشجيرات محضّرة مخبرياً بأنه عندما يكون لدينا جينات مضاعفة، فستخصص كل نسخة وظائفها. كمثال، يكون الجين adhA مختص باصطناع الأنزيم alcohol deshidrogenasa A. يعبّر هذا الجين في كل الانسجة النمائيّة في النبات، مع هذا تهتم نسخة الجينوم A بالتعبير في الجذور والفلقات، بينما تهتم نسخة الجينوم D بالتعبير في الاوراق والقِنابات { القِنابة: هي ورقة في قاعدة الزهرة أو ساقها .. قاموس المورد اسباني عربي }. ليس هذا فقط، كذلك لا تُعبّر النسخة A للجين adhA لا في التويجات ولا في الاسدية {جمع السداة: العضو الذكري في الزهرة }، لكنها تعبّر في النسخة D، من جانبها، في الجزء الأنثوي من الزهرة carpelos تعبّر فقط في النسخة A، وليس في النسخة D. تعبيرات مختلفة للجين بشجيرات القطن.تشكل الاشرطة الجينات المعبرة في كل نسيج،بصيغة أنّ At يكون نسخة A و Dt يكون نسخة D ، A جين adhA ، B جين adhD، Cجين A1520، D جين B5 لا تشكّل التباعدات في نماذج التعبير تلك حالة مُنفردة، بل تشكّل القاعدة في الجينات المُضاعفة لنوع Gossypium. دون الذهاب بعيداً، تسود في الجين adhD النسخة A في كل الانسجة عدا الأسدية، حيث تسود النسخة D . بالنسبة للبروتين A 1520 فالنسخة D هي التي تعبر باغلبية الانسجة الإنباتيّة، بينما تعمل النسخة A على اجهزة التكاثر. من جانبه، بالنسبة للجين B5 المسؤول عن الانزيم enzima oxalato oxidasa، فالنسخة D لا تعبّر في الاسدية، فهذا شيء تقوم به النسخة A. تُبيّن نتائج ابحاث اخرى، اضافة لكل ما سلف بأنّ كل تلك التعديلات في نماذج التعبير يمكن اعطاؤها مباشرة إثر عملية التضاعف الجيني. تبيّن تلك التفاصيل حالة جميلة للتطور الجزيئيّ، حيث نرى بأنه متى تضاعفت الجينات، ستتطور بصيغة مستقلة وتقوم كل نسخة متخصصة بنشاطها حول بعض الانسجة أو بعضها الآخر. الشعبة الجينية لنوع شجيرة القطن Gossypium sp لكن ايضاً التدجين، قد امتلك نتائجه يكون تطور القطن خلال عملية التدجين مرئياً في بنية تلك النباتات البيضاء السللوزية التي تحيط ببذور، والتي نوظفها بصنع الانسجة نحن. لا تكون الياف القطن شيء مُختلف عن خلايا أدميّة معدّلة تسمى tricomas، والتي تقوم بإحاطة البذرة. في غالبية انواع شجيرات القطن، تكون تلك الالياف كثيفة صغيرة جداً { أقل من 5 ميلمتر }، سميكة ومتلاصقة بشدّة، غير مفيدة كالياف نسيجية كما هي، حيث يصعب التفكير بأنها شجيرات قطن اصلاً. ففي الحالة البريّة، فقط شجيرات القطن في المجموعتين A و AD يُنتجان الياف اكثر طولاً بقليل ورخوة بحيث يمكن تحويلها الى خيطان مفيدة، وهذا ما دفع الانسان للاستفادة منها، حيث يكون النوع المنتمي للمجموعة AD الافضل كنوعية. ما قبل – التكيُّف هذا { المؤكد أنه بسبب الانتقاء بعلم تشكُّله " بمورفولوجيته " بوصفه " ناثر " للبذور }، سيسمح للانسان بالقيام بالانتقاء لتلك الانواع لاجل زراعتها. في الانواع المزروعة، تكون الالياف أكثر طولاً بكثير، قوية ورقيقة أكثر مما لدى النوع البري. نرى في حالة شجيرات القطن AD بأنها طوّرت توزيعاً في المهام لاجل تشكيل الالياف، حيث يتحمّل الجينوم A عبء نموها وتطورها، بينما يتحمّل الجينوم D عبء نوعيتها ونظافتها. من جانب آخر، شجّع التدجين في شجيرات القطن الزراعية القديمة في المجموعة grupo A على تطور الالياف المتشابهة كثيراً مع ما تمتلكه العناصر المزروعه من المجموعه grupo AD، وقفاً على تأثير ضغط انتقائيّ شديد الشبه. تدجين شجيرات القطن نوع (Gossypium sp) لكن الافضل هو ما يحفظه علم الجينات المُقارن لعناصر المجموعة grupo A، التي تُنتج الياف مفيدة مع مجموعات اخرى من شجيرات القطن الغير قادرة على انتاج تلك الالياف. على الرغم أنّ تكوين الالياف، يستلزم عملية معقدة يتدخّل فيها مئات الجينات ومسارات تعبير، يكون جوهرياً ما يحدث خلال اللحظات الأُوَلْ. نجد في شجيرات القطن تلك التي لا تُعطي اليافا مفيدة، بأن ذاك الانخفاض بنمو اليافها، يحدث بسبب تحرّر عوامل أكسدة واضطراب خليوي { الماء المؤكسج H2O2وأنواع مفعّلة من الاوكسجين ROS }، وهذا يؤدي الى اعاقة نمو الالياف. لا يحصل ذات الامر في المجموعه grupo A. فتمتلك كل شجيرات القطن جينات عديدة الوظائف، حيث نجد: يتحمل الجين GAST1 مسؤولية تنظيم اشياء بينها الانقسام والنمو الخليوي، فيما يساهم الجين Cop 1 في تنظيم الموت الخليوي، ويقوم الجين Pex1 بتنظيم تكوُّن peroxisomas . تُعلّم الدراسات الجزيئية بأنّ كل تلك الجينات قد تجندَّت خلال تطور المجموعه grupo A لاجل تخفيض العناصر المؤكسدة خلال نموّ الالياف، بصيغة تسمح بإطالة فترة نموها وبالتالي اعطاء الياف مفيدة من شجيرات قطن بريّة في المجموعتين grupos A و AD . لكن تلك الالياف لا تزال تعاني القزامة. فالقطن الذي نعرفه هوثمرة ضغط انتقاء قد أسسه التدجين البشري. هكذا تمتلك شجيرات قطننا الحديثعناصر ضابطة تمارس فعلاً أكثر قوّة، الامر الذي يقود في تركيزات خفيضةجداً لانواع مفعّلة من الاوكسجين (ROS) خلال زمن اطول بكثير، فتكون نتيجته نموّ مهم بطول الياف القطن الذي نتمتع باستعماله بيومنا هذا. لاحظ الباحثون ايضاً بأن عملية تطور الاستقلاب المديد في الزمن، قد ظهر بشكل متزامن مع تدجين هذه النبتة. نماذج مقارنة لتطور ونمو الياف شجيرات القطن. التدجين في المكان المعروف باسم العالم القديم، المتضمن اوروبا، افريقيا وآسيا. كان النوع الاكثر اهمية تقليدياً هو G. herbaceum. الذي يُظن بأنه نشأ في جنوب افريقيا. مع ذلك تتركّز الزراعات الاقدم في الشواطيء الهندية. ليس غريباً ان يعتبر بعض الباحثين الهند كمنطقة انتشرت منها بذور هذا النوع. فاقدم الاشارات لاستخدام القطن في انسجة وحبال تعود الى ما قبل 4300 – 3750 عام في الهند وباكستان، ففي طبقات موقع yacimiento de Harappa آثار أخرى لاستخدام نوع القطن G. herbaceum في اماكن قريبة. خارجة انتشار وتوزّع نوع قطن Gossypium herbaceum L تُشير الاسماء لاصول مختلفة. حصل وصول القطن لمناطق المتوسط بوقت متأخر أكثر. من اوائل الاشارات الى القطن، تلك التي جاءت بوثائق آشورية تُخبر عن " اشجار تُعطي صوف " والتي زرعها الملك سنحاريب، ملك آشورية Asiria. تعود تلك الوثائق الى 694 قبل الميلاد، مع هذا فالاشارة الى صيغة شجرية تدفع للشكّ بأنّ النوع المُدرج هو G. arboreum. أما وثائق الميشنا والتلمود اليهودي فتشير الى القطن، ما يعني احتمال زراعته في القرنين السادس والخامس قبل الميلاد في شرق المتوسط. يبلغ الشرح الوافي عن زراعة القطن ذروته مع اوائل العصور الاسلامية، بنهايات القرن السابع الميلادي.  لوحة منشورة في الفترة 1357-1371 من قبل John de Mandeville لرسم شجيرة القطن على شكل خروف كرمز لنوعيّة ما تعطيه ثمرة القطن! خيال خصب! تختلف قصة انواع العالم الجديد، خصوصاً، نوع G. hirsutum، حيث تمتلك هذه النبتة معدل انتشار طبيعي في قسم كبير من القارة الاميركية والكاريبي، عبر تدرُّج تشكيليّ من الانواع البرية المختلفة الى الانواع المزروعه. فنجد اسلاف في وادي Tehuacán تعود الى 4000 - 5000 عام قبل الآن، مع ذلك، تحضر البقايا الاثرية الاقدم المعثور عليها في كهف Ocampo في المكسيك بعمر 5800 عام. من غير المُستبعد بأنّ تدجينه سيبدأ في شبه جزيرة Yucatán مع هذا، بدأ فقط اعتباراً من القرنين التاسع عشر والعشرين استثمار هذا النوع الهام من القطن بمستوى عالميّ. خارطة انتشار وتوزّع نوع القطن Gossypium hirsutum L تُشير الاسماء لاصول مختلفة  نوع Gossypium darwinii أو قطن داروينالمستوطن في جزر غالاباغوس Galápagos يتبع |
|
|
|
|
|
|
08-06-2016, 08:48 PM
|
رقم الموضوع : [6] |
|
عضو ذهبي
|
البُستانُ التطوريُّ {6}: تطوُّر الشعير يفقد عقله دون تلفزيون ودون بيرة!! شخصيّة رسم كارتوني شهير Homer Simpson الجعة أو البيرة. ذهبيّة وساحرة. سوداء وذات رغوة. في عبوات متنوعة وكؤوس مختلفة. هل يتخيل احدكم عالماً دون بيرة؟ بماذا نسترطب في الصيف؟ بأيّ شيء نصحب بعض المازوات وبعض الموالح؟ كيف يعيش الاجانب الذين يقصدون سواحلنا للسياحة؟ مَنْ يحفظ سهراتنا؟ ماذا سيحلّ بإيرلندا؟ تكون المادة الاساسية بالبيرة حبيبات الشعير، التي تفيد بتحضير انواع خبز وبسكويت مختلفة ومنتجات اخرى. كما ان الشعير يدخل بتحضير اعلاف للخنازير والابقار وغيرها من الحيوانات الداجنة. بالتالي يمكننا القول بأن البيرة والشعير: كانا معنا منذ الازل. أو ربما لا؟ يشكّل الشعير نباتا هائل الانتشار. في وقتنا الراهن يأتي خلف الذرة، القمح والأرز، بوصفه رابع نبات حبوبيّ والاكثر اهمية على الصعيد العالمي. يكون اسمه العلمي هو: Hordeum vulgare ssp. vulgare L يوجد جنبا إلى جنب مع القمح، الذرة، الأرز، الذرة البيضاء، قصب السكر أو البشتة { نبات سنوي من النجيليات يُزرع للاستفاده من عشبه .. قاموس المورد اسباني عربي }. يكون الشعير نبات سنوي، تكون طبيعة جينومه ثنائيّة diploide { كجينومنا االبشري }، يحتوي على 7 ازواج من الكروموزومات فقط. وبعكس نباتات اخرى، يمتلك مستوى مرتفع من التلقيح الذاتيّ، ما يعني كون التلقيح التهجيني نادر جداً، هكذا تكون ازهاره ذاتها التي تتكاثر مولدة ذاتها. سنبلة ناضجة من شعير نوع Hordeum vulgare ssp. Vulgare يشكّل الشعير بوقتنا الراهن نبات سهل الزراعة والحصاد، متكيف مع كل انواع البيئات والمناخات، حيث يتوزع بكل انحاء العالم. مع هذا، في زمن آخر، في مكان مفقود، كان الوضع مختلف بشكل ملموس. انتاج العالم من الشعير لكل هكتارفي الفترة 2000 – 2002 بدايات قديمة، ارتباطات شديدة منذ 23000 عام، بلغت الحقبة الجليدية أوجها. بينما تراجعت في نصف الكرة الشمالي، في الهلال الخصيب حيث تراوحت درجات الحرارة بين 12 و16 درجة مئوية كمتوسط سنوي، وهو متوسط أقل بست درجات من متوسط العصر الحديث. كما كانت الهطولات المطرية أقلّ، حوالي 300 الى 450 مليمتر سنوياً، مقابل 500 مليمتر يمكنه " التمتع بها " حالياً. تحت وقع هذا المناخ الرطب والصحراوي، نجد ملاجيء غنيّة بالحياة النباتية قادرة على دعم العُطم { جماعة الحيوانات والكائنات الحية في منطقة او حقبة جيولوجية .. قاموس المورد اسباني عربي } المتكونة من الغزلان، القوارض والطيور، بينما تسبح اسماك ورخويات في بحيرات قريبة. وفي قمة الهرم الغذائيّ لدينا الجماعات البشرية المكونة من مُلتقطي الثمار والصيادين. ماكيت لمكان اثري اسموه Ohalo II في حيفا في فلسطين والذي يعود الى قبل 23.000 عام هذا يوفر لنا احد افضل الطبقات الآركيولوجية في العالم، موقع Ohalo II الواقع في فلسطين الحالية على ضفاف بحيرة طبرية أو بحيرة الجليل، عباة عن بلدة قديمة محفوظة بشكل جيد والتي تعود لما قبل 23000 عام من القدم، عندما بلغت الحقبة الجليدية الاخيرة خواتيمها. كانت الحفريات بقيادة Daniel Nadel من جامعة حيفا، بين العامين 1989 و1991 وبين العامين 1998 و2001، عندما انخفض مستوى بحيرة طبريا بشكل كافٍ، الامر الذي سمح بالحفر بتلك المنطقة. تمثيل لموقع Ohalo II في معرض بمتحف Hetcht جامعة حيفا .. فلسطين تبدو البلدة كمخيّم. فالخيم الستة كانت مصنوعه من الاغصان والاوراق، والقبر المجاور احتفظ بجسد. لكن أكثر ما لفت الانتباه هو الادوات المعثور عليها. لقد كانت معالجة المادة النباتية لاجل التغذية عادة قديمة جداً، فلقد احتوت الادوات على طواحين حجرية { مفترض انها مستعملة لطحن الحبوب } في العصر الحجري الاقدم { بين 45000 و18000 عام }. مع ذلك، عثروا في الموقع على ادلة مباشرة لشيء اكثر تعقيد، تحميص الحبوب والفواكه قبل معالجتها. عثروا على احجار كربونية مُحضّرة جيداً أكثر او اقلّ. وفي المحيط حبيبات معرّضة للحرارة. يصعب تخيُّل كونها شيء آخر مختلف عن مواقد بدائيّة، وتُشكّل الدليل الاول على تحميص الفواكه والحبوب لاجل التغذية. وهذا لا يكون هزلياً. فربما بالنسبة لاولئك الناس سهّلت المحمصات الاجراءات وزادت الاستساغة مع هذا، بصورة غير واعية، بفضل المحمصات ايضاً: كانت تزداد القيمة الغذائية والقدرة الهضمية. كانت الطبقة محفوظة جيداً، وكان بالامكان دراسة النظام الغذائي لاولئك الناس. وبالتأكيد كان نظام متنوع ينطوي على فواكه وبذور لكثير من الانواع المتواجدة في المحيط، مثل البلّوط Quercus ithaburensis، اللوز، التين البري، الفستق الحلبي، الزيتون البري، ثمار شوكية Ziziphus spina-christi، العليق، العنب البري....الخ. مع ذلك، كان المورد النباتي الاكثر اهمية هو النجيليات. كانت ازمنة صعبة والموارد محدودة. لا يكون غريباً أن كثير من الاعشاب التي تُعتبر بيومنا هذا " اعشاب ضارة "، كانت في تلك الحقبة ذات اهمية بالغة. ففي الموقع المذكور Ohalo II كان هناك انواع " ذات حبيبات صغيرة " مثّلت ما نسبته 34.6% من الحجم الكامل للنجيليات الراهنة المستعملة في التغذية. انتمى القسم الباقي الذي نسبته 65.4% الى الحبوبيات، بالتأكيد الى " الشعير البري " Hordeum vulgare ssp. spontaneum وإلى " النشوي البري " Triticum turgidum ssp. dicoccoides والتي تكون اسلاف الشعير والقمح الحديثان. لكنها لم تكن تحتوي على اجزاء متماثلة. كلا، فلقد تجاوز الشعير للنشويّ بنسبة 25:1 . جدول يبيّن انواع عديدة من النجيليات استخدمها بالتغذية سكان موقع Ohalo II ، تمثل الارقام كميات البذورلا احجامها. ترمز SGG كمختصر كلمات انكليزية Small-Grained Grasses ، أي اعشاب ذات حبيبات صغيرة ولادة عصر جديد بمضي الزمن، ازدادت اهمية الحبوب دون توقف، على حساب تلك " الاعشاب ذات الحبيبات الصغيرة " التي كانت تفقد اهميتها تدريجياً لتصل لمرحلة الاختفاء. جدول يوضح تطور اهمية الحبوب في التغذية. . ترمز SGG كمختصر كلمات انكليزية Small-Grained Grasses ، أي اعشاب ذات حبيبات صغيرة اسم الشعير البري العلمي هو: Hordeum vulgare ssp. spontaneum سيكون أمّ شعيرنا الراهن: Hordeum vulgare ssp. vulgare L. وهي نباتات نعتبرها اليوم كانواع فرعيّة، ما يعني، توجد في طريق انفصال لانواع مختلفة. يكون " الشعير البري " عبارة عن عشبة سنوية متكيّفة، نعثر عليها في مناطق ذات مناخ متوسطي { شتاء ماطر وصيف جاف } وفي الصحاري حيث يندر المطر، نجدها على مستوى سطح البحر وفي ارتفاعات تصل الى 1500 متر عن سطح البحر، تغزو قطاع بطول اكثر من 3500 كيلومتر من شواطي الشرق الاوسط المتوسطية الى جنوب شرق آسيا، بما فيها سلسلة جبال الهيمالايا والتيبيت. نماذج من الشعير البري Hordeum vulgare ssp. Spontaneum منذ 15000 عام، وصلت جماعة بشرية اخرى إلى الهلال الخصيب، عُرفت باسم Natufians، وظهر أفرادها لاول مرة كجماعة صيد – التقاط ثمار، مع هذا، خلال فترة قصيرة استقروا على شكل بلدات نصف – مستقرة على اراض خصيبة في شرق المتوسط حيث يتوفر الغذاء بصيغة اغذية نباتية متوفرة بكثرة، وسيكون الأمر افضل من الترحال الذي نتج بوصفه غير ذي جدوى بعد الاستقرار. توجّب عليهم معرفة التعامل والتحكّم بهذا الغنى الطبيعي الوافر. أتقنوا صنع طواحين الحجر المفيدة بتسهيل طحن الحبوب لاجل استخراج الاصباغ، بدؤوا بصنع المناجل الحجرية لاجل تقطيع العشب، هكذا كما المواقد والحفر لاجل تحميص الحبوب. كذلك طوّروا تقنية التخزين، والتي تركزت وقتها في آبار متواضعه محفورة بالارض، حيث كان بالامكان مراكمة الغذاء في موسم فيه فائض انتاج. أطلق بعض الباحثين على هذا " ما قبل التكيُّف ". قياساً بتطورات ستحصل في مستقبل بعيد، يتميز " ما قبل التكيُّف " بتطوير ادوات تسبق تطبيقات لا يمكن الاستغناء عنها فيها، هنا نتحدث عن التقنيات الضرورية بادارة المواد النباتية قبل ظهور الزراعة بكثير. رسم تمثيلي لجماعة من Natufians، بفترة ما قبل 11.000-15.000 عام بحصول تغيُّر مُناخي قبل 12800 عام، حدثت حقبة جفاف لعدّة عقود معروفة تحت اسم: Younger Dryas، حيث ارتفعت درجة الحرارة من 5 الى 10 درجات مئوية وهو ما ساهم بانطلاق الزراعة. يسعى بعض الباحثين للربط بين كلا الحدثين للبرهنة على أنّ جماعات محددة Natufians راكمت خبرة على مدار آلاف الاعوام في معالجة وتخزين انواع نباتيّة، و كردّ من تلك الجماعات على نقص الانتاج الطبيعي الناتج عن التغيُّر المُناخي المُشار له سالفاً، قاموا هم ذاتهم بزراعة الارض. بكل الاحوال، عندما بدأت الزراعة، كان الشعير من النباتات الرائدة. تعود اوائل مؤشرات تدجينه الغير قابلة للنقاش للفترة الممتدة بين ما قبل 10.500 الى 10.000 عام في منطقة الهلال الخصيب Creciente Fértil. وهي منطقة واقعة في الشرق القريب جنوب هضبة الاناضول بين نهري دجلة والفرات. إثر 2000 عام من الزراعة، اي منذ 8000 عام، بدأت زراعة الشعير بالانتشار نحو شمال افريقيا واوروبا. التوزُّع الجغرافي لاهم الانواع البريّة من الشعير لكن يوجد مناطق تدجين اخرى، حيث كان " الشعير البري " واسع الانتشار من شواطيء البحر المتوسط الى مركز قارة آسيا. وباستكمال الدراسات الوراثية، يتم اعتبار الحقبة الواقعة بين 10000 الى 8000 عام قبل الآن: هي التي حصل فيها ابتكار الزراعة المستقلة للشعير في جنوب غرب آسيا، كما تُشير طبقات وادي Indo وفي Megrah { قبل 8000 عام }، أو في Jeitun { قبل 7000 عام } في المنطقة الشمالية الغربية من الهضبة الايرانية وجنوب تركمانستان. تدجين الهلال الخصيب، كما تدجين مركز آسيا: قاما بتطوير الشعير بصورة متشابهة. تكيّفت كلا السلالتين بشكل متوازي ومتطابق الميزات تقريباً. كان هناك حتمية بالتغيُّر في الشعير. بالنسبة للنبات، يمثل التدجين صيغة مثالية لمضاعفة نجاحه التكاثريّ وحجم رقعة انتشاره. لكن هذا ليس دون ثمن أو مجّاني، فمن بين أكثر الضرائب التي عليه دفعها: حريته في الانتشار { المقصود هنها بالطبع الاحتكار الزراعي التجاري الصناعي للنبات وفائدته لبلد المنشأ ودخوله في التنافس .... فينيق }.  سنبلة مزهرة للشعير البري Hordeum vulgare ssp. Spontaneum ضريبة التبعيّة تشكّل السنيبلة نظام الازهرار الاساسيّ في النجيليات. تتكوّن من زهرة أو اكثر، حيث تستند كل زهرة على وريقتين او ثلاث وريقات صغيرة الحجم تُدعى lodículas. بدورها كل زهرة مع lodículas تكون مغطاة بمصراعين محددين باسم lema واسم pálea على التوالي. تتحد تلك الاخيرة بالنهاية لتشكل ساق ينقسم الى شقين brácteas متعارضين يسميان glumas، تكون كل هذه البنية للآن هي السنيبلة. وفي حال كون السنيبلة هكذا، فما هي السنبلة اذاً؟ تكون السنبلة ببساطة عبارة عن تجمُّع من السنيبلات لا اكثر ولا اقلّ. في اليسار تمثيل تشريحي لسنبلة نوع Avena sterilis ، في اليمين سنيبلة لجنس Avena ملاحظة: تشكل brácteas اعضاء نباتية تذكرنا بالاوراق، لكنها الاوراق الواقعة في جوار الازهار. أن تستخدمنا نبتة لاجل نشرها، عبارة عن ابتكار حديث جداً. فحتى ظهور الزراعة، اكتفت كل انواع النجيليات بذاتها لنشر بذورها. لهذا تضم سنابل وسنيبلات متكيفة بشكل كامل للانتشار بواسطة الرياح anemocoria و / أو بوساطة الحيوانات zoocoria . واسلاف نباتاتنا المزروعة لم تكن أقلّ. مثل: النشوي البري Triticum turgidum ssp. dicoccoides كسلف بري للقمح، يمتلك الشعير البري Hordeum vulgare ssp spontaneum سنبلة هشّة، متى بلغت سنّ البلوغ تنكسر بسهولة محرّرة السنيبلات. والتي بدورها تتكون من وريقات تنتهي بسفاة { شعيرات السنبلة } طويلة مزوّدة بما يُشبه الكلّابات الصغيرة مفيدة لاجل التعلّق بشعر مواليد ثدييات، سواء ماعز، أيّل، خنزير بري أو غزال. { لاجل انتشار بذورها بواسطة تلك الحيوانات حين تعلق بها وهنا تكمن الفائدة .. فينيق } سنيبلات الشعير البري نوع Hordeum vulgare ssp. spontaneum ، لاحظوا الشعرات الحاضرة في سفاة الوريقات شيء مشابه لما نراه كل يوم في نوع Hordeum murinum L والمعروف تحت اسم " شعير الفئران ". يمتلك قريب الشعير هذا العادة السيئة بتعلّق سنيبلاته بكلساتنا { جراباتنا }. لا يمتلك هذا النوع كلّابات في السفاة، بل في قاعدة السنيبلة ذاتها. هذا مثال عن دور الحيوانات بنقل البذور.في هذه الحال، تنتقلسنيبلات نوع Hordeum murinum عبر البشر! يكون عالم الاحياء الياباني Takao Komatsuda، باحث منذ العام 1985 في المعهد الوطني للعلوم الزراعية الحيوية Tsukuba في اليابان، واحد من الخبراء الحاليين في علم وراثة وتطور الشعير بطول فترة تدجينه. من بين ابحاثه ما يتصل بالجينان Btr1 و Btr2. حيث يكونان مسؤولان عن تكوين سنبلة هشّة تنتشر بواسطة الحيوانات. مع ذلك يتزود الجينان بزوج من الأليلات ذات طبيعة متنحيّة، هما: btr1 و btr2 واللذان يحثّان على نمو سنابل غير هشّة. يكون كلا الاليلين btr1 و btr2 نادرين في الشعير البري. مع هذا فقد حولهما فعل التدجين الانتقائي الى صيغة الشعير الزراعي مغيّراً الى الابد طبيعة سنبلتها. وهذا لم يحصل بمرّة واحدة فقط، بل بمرتين: يسود الأليل btr1 في الزراعات الغربية اكثر، بينما يسود الأليل btr2 في الزراعات الشرقية اكثر، دليل اضافي على الاصل الثنائيّ لزراعة هذا النبات. من جانب آخر، نعرف بفضل علم الآثار بأنّ ظهور السنابل الغير هشّة قد كان عملية تدريجية. فقد مثّلت هذه الميزة أكثر من 30% من اجمالي النوع، منذ الفترة الواقعة بين ما قبل 9300 – 8500 عام، لترتفع نسبتها الى 60% في الالف عام التالية، بالتالي، السنبلة غير الهشّة المنغلقة indehiscente، قد شكّلت جزء من الشعير بطرق التدجين.  Aسنبلة حديثة هشّة لنشويّ بريّ، Bسنيبلة، Cاجزاء السنيبلة بالتفصيل، D قمح مدجّن غير هش، E تفصيل لانكسار غير مُنتظم للسنيبلة، F الرسم يبيّن التعاقبات المختلفة لسنيبلات السنابل " الهشّة " باللون الاخضر الغامق،ولسنابل " غير هشّة "باللون التوتي، حيث ينتمي موقع الأسود Aswad ورماد Ramad لحالات فيها شعير زيادة الإنتاج {1} أثّر التدجين ايضاً في بنية السنبلة. في الشعير، تتكوّن السنابل من ثلاثة سنيبلات { واحدة في المركز واثنتان في الطرفين } مشدودة الى محور طويل. بناء عليه، نقوم بالتمييز بين نوعين رئيسيين للسنبلة ذات 6 صفوف، حيث تكون كل السنيبلات خصيبة، وذات صفين، حيث تكون السنيبلة المركزية خصيبة فقط بينما السنيبلتان الطرفيتان منخفضة النمو ومصابة بالعقم. وفق النموذج الاكثر قبولاً، فقد تولّدت تنوعات الصفوف الستّة اعتباراً من تنوعات الصفوف الثنائيّة. دون الذهاب بعيداً، يمتلك نوع H. vulgare ssp. Spontaneum سنابل ذات صفين، وهو تكيُّف يسمح لسنيبلاته بالتدحرج على الصخور والاحجار عند الوقوع على الارض. مع هذا، في مناطق محددة وسط آسيا، تمّ العثور على تنوعات من " الشعير البري" ذو 6 صفوف والمسمى H. vulgare ssp. Agriocrithon والذي كان مختلف بشكل واضح عن هذا النموذج. هل يمكن ان يشكّل نوع H. vulgare ssp. Agriocrithon السلف الحقيقي لشعيرنا المزروع؟ أو انها تشكّل حالة استثنائية؟ ما هو اصلها؟ بيومنا هذا، يكون مقبولاً القول بأنّ هذا النبات: لا يكون سوى نتيجة التهجين بين الشعير المزروع والشعير البري. في الاعلى سنبلة شعير بري،في الوسط سنبلة شعير " ذات صفين" ، في الاسفل سنبلة شعير " ذات 6 صفوف " سنتكلم الآن عن الدليل الاثري المتوفر. حتى قبل 9000 عام من الآن: كان الشعير ذو صفين غير قابل للتمييز عملياً عن " الشعير البري ". حتى ان السنابل قد احتفظت بخاصيتها تلك ببداية التدجين. فقط في طبقات تعود الى الفترة 8800-8000 عام قبل الآن، نبدأ بملاحظة حضور بقايا تنوعات مزروعة منه { يمكن تمييزها بفضل سنيبلاتها المنغلقة } ذات الست صفوف، والتي ستسود في الاراضي الخصبة، في طمي الهلال الخصيب وبلاد الرافدين منه على وجه الخصوص كما في مصر القديمة منذ 7000 و 6000 عام، حيث جرى استبدال السنبلة ذات الصفين واعتماده كزراعة ذات اهمية خاصة بحضارات العصر الحجري الحديث Neolíticas في شرق المتوسط. عملة من مستعمرة يونانية قديمة في ايطاليا، في الفترة 330-340 قبل الميلاد، كانعلى أحد وجهيها الشعير ذو الست صفوف. ايضا يوجد في وجه قطعة العملة الآخر Leucippus كأول يوناني طوّر الفكرة القائلة بأنّ المادة مكونة من ذرات بالنهاية، نتابع مع دراسات جزيئية مُتواصله مع العالم Takao Komatsuda. يكون عدد الصفوف مضبوطاً بواسطة جين وحيد هو Vrs1. والذي نعثر على أليلين له، واحد سائد (Vrs1) مسؤول عن خاصية " الصفين "، وآخر متنحي (vrs1) مسؤول عن خاصية " الست صفوف ". عمل الجين لا يكون صعباً على الفهم. ينتمي هذا الجين لمجموعة بروتينات تسمى: IHD-ZIP، هي بروتينات قادرة على التعرُّف على سلاسل مؤكدة من DNA والاتحاد بها مانعة أو مفضّلة { بحسب الحالة } التعبير لمجموعات محددة من الجينات، هكذا يقوم الأليل Vrs1 بتركيب بروتين، والذي باتحاده بجينات محددة، يكبح نمو وبلوغ السنيبلات الجانبية في السنابل ثلاثية السنيبلات. سبّبت طفرة في الأليل Vrs1 بتوليد أليل vrs1، الذي يقوم بتركيب بروتين لا يمكنه القيام بوظيفة كتلك. بصيغة لا يمكن فيها لايّ كان تحقيق منع او كبح نمو السنيبلات الجانبية، حيث انها تنمو معطية المكان لظهور سنيبلات ذات 6 صفوف. هذا حدث بصيغة مستقلة على الاقل في ثلاث مناسبات: في غرب المتوسط، في شرق آسيا، ويكون اليوم الأليل الناتج ذو توزُّع عالمي. اختفى نوع الشعير ذو الصفين من الهلال الخصيب وشمال افريقيا منذ 7000 – 6000 عام، ليعود ليظهر كما لو انه من اللاشيء في هاتيك المناطق. حدث هذا حوالي القرن العاشر الميلادي، إثر هذا بقرنين او ثلاثة قرون، ربما جُلِبَ من قبل الحملات الصليبية، حيث وصلت تنوعات الشعير ذو الصفين إلى اوروبا، وساد بعد ذلك تنوّع الشعير ذو الصفوف الستّة. تدرُّج بسنيبلات ذات صفين وذات 6 صفوف، A تنوعات أثيوبية، B شعير برّي، C نوع مزروع " ذو صفين " من السنيبلات الجانبية العقيمة، D شعير بري نوع proskowetzii، Eشعير مزروع " ذو 6 صفوف " بذور عارية كما رأينا في المواضيع السابقة، فإنّ بذور اسلاف الذرة والقمح محميّة بغطاء نباتي، حماية قد اختفت عند متحدريها المزروعة كنتيجة لانتقاء مفروض خلال عملية التدجين. مع هذا، يكون الوضع مختلف في شعيرنا، فبذوره محميّة بغطاء ملتصق بشدّة بها، حيث يتكوّن من lema و pálea . حسنا الآن، كيف تمّ الاحتفاظ بهذه الحماية؟ يكون السبب في استعمالات النبات: فاغطية كتلك تنتج بوصفها مفيدة لاجل معالجة المُلت { الحبوب الناتشة صناعياً ولا سيما حبوب الشعير التي تستعمل في صناعة الجعة او البيرة .. قاموس المورد اسباني عربي } خلال عملية تحضير البيرة، من جانب آخر، يُستخدم الشعير كغذاء للحيوانات التي لا تشتكي عادة عندما تصادف بذور محمية بهذا الشكل. من يشتكي بالغالب هم البشر. انواع الشعير التي نستعملها بتحضير منتجات المعجنات نعم تكون بذورها عارية، ومن المعتاد ان تكون ذات 6 صفوف وقد ظهرت لاول مرة منذ 8000 عام، قبل ان تنتشر زراعة الشعير من الهلال الخصيب والهند الغربية الى باقي العالم القديم. عري تلك البذور يكون مشروطاً بوجود جين وحيد اسمه Nud، يقوم بتركيب بروتين يضبط تخزين سلسلة من الشحميد على سطح البذرة، والتي تظهر بكونها اساسية لاجل السماح بالتصاق وثبات lema و pálea عليها ذاتها. لا تسمح طفرات هذا الجين بتخزين شحميد، ولا يمكن التصاق تلك الاغشية النباتية، ما يحقق استمرار العُري للبذرة. بالحال تمّ انتقاء تلك الطفرات لتصبح تنوعاً مالوفاً لكثير من الانواع المزروعة من الشعير. في انواع شعير " ذو صفين "، A الى اليساربذور مغطاةB .الى اليمينبذور عارية، سنابل idem زيادة الإنتاج {2} بلغت سنبلة الشعير الكمال في الفترة { 10500-9500 عام قبل الآن }، لتكون مضاعفة ب3 لعدد السنيبلات التي كانت قادرة على الانتاج في الفترة { 8800-8000 عام قبل الآن } وفقدت تغطيتها الحمائية " التي صعّبت " التحكم بها كغذاء في الفترة { 8000 عام قبل الآن }، ايضاً كان حجم حبّتها آخذ بالنموّ. مقطع عرضي بحبيبة شعير بري في اليسار، ولنوع مزروع في اليمين في هذا الجانب، تكون السجلات الاحفورية وصفيّة بقوّة. فقد قام الباحث الاثري النباتي George Willcox العامل في المركز الوطني للبحث العلمي في ليون – فرنسا بمقارنة بذور الشعير البري { في عملية تدجين واسعة } بطبقات أثرية مختلفة في حوض الفرات الواقع شمال الجمهورية السورية، في منطقة جرف الاحمر الباكر { 9800-9700 عام قبل الآن } والمتأخر { 9400-9300 عام قبل الآن }، وموقع دجادي { 9400-9000 عام قبل الآن } وكوزاك الشمالي { 6050 ± 100 عام قبل الآن }، حيث يُعتبر الموقع الاخير تمثيل غير مسبوق لمرحلة مسماة Calcolítico، عندما بدأ البشر لأوّل مرة باستعمال المعادن. تبيّن دراسة Willcox اتجاهاً هاماً، ولو انه بطيء وتدريجي زمنياً، نمت خلاله بذور الشعير في الحجم كما في الثخانة. مقارنة بين بذور الشعير في المواقع المذكورة اعلاه في حوض الفرات. لاحظوا النمو في الثخانة، احداثيات رأسيه، وفي العرض، احداثيات افقية، بمضي الزمن وهذه لم تكن كل التغيرات الحاصلة. فتحتاج كثير من تنوعات القمح والشعير، كما اسلافها البرية، إلى بضع اسابيع من التعرُّض للبرد { استبراد } – بين 0 الى 10 درجات مئوية – لكي تتمكن من الازهرار بشكل طبيعي، ولن يتحقق الإزهرار دون هذا التعرُّض للبرد. هذا الاحتياج التجلُّدي، اسمه: الاستبراد { معالجة البذور بالبرد بغية تسريع انتاشها – انباتها – وتسريع نموّ النبات وأزهاره .. قاموس المورد اسباني عربي }. هكذا تسمى هذه النباتات: بالنباتات الشتوية، تُزرع بأواخر الصيف أو بدايات الخريف، حيث تتعرّض للبرد خلال الشتاء، وتُزهر بقدوم الربيع. لكن البرد الزائد يقضي على الغرسات، اي باماكن اكثر ارتفاعاً، مع هذا يُزرع في تلك الارتفاعات الشعير والقمح، لكن كيف يمكن تحقيق هذا؟ يشكّل الاستبراد عملية معقدة، يشترك فيها عدد من الجينات، يكون اهمها هذه الايام: اولاً: VRN1، يكون تركيبه مُحفزّاً من البرد، يقوم بكبح الجين VRN2 ويحثّ على تحقيق الإزهرار. ثانياً: FT، عندما تكون الأيام طويلة { مدة النهار طويلة }، يقوم بتحفيز تركيب الجين VRN1 وتسريع. ثالثاً: VRN2، يقوم بكبح الجين FT وانهاء الإزهرار. علم وراثة استبراد البقوليات هكذا يقترح نموذج راهن، بأنّه خلال فترة شباب النبات، أثناء الصيف و الخريف: يمنع الجين VRN2 تعبير الجين FT، وباعتبار أنّ مستويات الجين VRN1 تكون منخفضة فلن يوجد اي حافز جيني لتحقيق الإزهرار. عند حلول فصل الشتاء، يقوم البرد بتحفيز تعبير الجين VRN1 الذي بدوره يقوم بإعاقة وتخفيض تركيب الجين VRN2. وحين حلول فصل الربيع، تكون مستويات الجين VRN2 منخفضة كثيراً، ما يجعل بامكان الجين FT التعبير بحريّة، الأمر الذي يحفّز تعبير أكبر للجين VRN1 ليبلغ تركيزاً عالياً فيمتلك " القوة " الآن لاجل تحقيق عملية الإزهرار. حسناً الآن تمنع طفرات ملموسة في الجين VRN1 ضبطه، فلزوم فترة البرد ليس لتعبير هذا الجين، بل لان تلك الطفرات سبّبت حدوث تعبير كذاك بصورة حادّة كل الوقت محفزة بهذا الشكل ازهرار سابق لأوانه. تصل طفرات اخرى في الجين VRN2 هذه المرّة، إلى منعه من تحقيق عمله، وكنتيجة لهذا يعبّر الجين FT الذي بدوره يحفز الجين VRN1، والذي بدوره يقوم بتحفيز الازهرار. جدول الضبط الجيني لاستبراد البقوليات في كلا الحالتين، لا تتطلب النباتات الاستبراد لاجل الازهرار. تحتاج تلك التنوعات من القمح والشعير لفصل الربيع، يمكن زراعتها بنجاح في مناطق باردة بهذا الفصل. حيث يمكن زرعها في بدايات الربيع ودون الحاجة لفترة باردة، بحيث تُزهر وتُثمر بحلول الصيف متفادية بهذا الشكل، الجليد الشتويّ الذي يقتلها. الجعة { البيرة }، متى؟ وأين؟ للآن رأينا متى عرفنا الشعير. اين ومتى بدأت زراعته. كذلك رأينا كثير من التغيرات المُرافقة لتدجينه، كالطفرة والانتقاء اللاحق، والقادرة على تحقيق تعديلات جوهرية في بيولوجية هذا الكائن. مع ذلك، ماذا كانت الجعة؟ أين وُجِدَ أصلها؟ لا تكون اسئلة سهلة. بحسب كثير من الباحثين، يعود تاريخها الى 8000 عام قبل الآن، في أوج تقنيات زراعية وبدايات صنع الخبز. فحبّة البقول بذاتها تكون ذات تغذية قليلة، فهي كربوهيدرات بشكل رئيسي مع احتواء بروتيني قليل { 12 – 13 % } ونقص كبير بالحموض الامينية والفيتامينات. مع ذلك، تتمكّن عمليّة التخمير من تبديل كل هذا. لوح مسماري يعود للعام السادس لتولي كاهن مدينة لغش السومري الحكم، قبل 2.370 عام.تكون وثيقة ادارية حول تحضير 3 انواع من الجعة وتبيان كميات الشعير اللازمة ومكونات اخرى ضرورية لتحضيرها تمّ تحقيق التخمُّر بفضل الخميرة، التي هي عبارة عن فطريات وحيدة الخليّة، ويشكّل النوع Saccharomyces الجنس الابرز منها، حيث تحوّل تلك الكائنات الحيّة في غياب الاوكسجين السكريات الموجودة في حبّات الشعير الى الايتانول { كحولنا } وثاني اوكسيد الكاربون (CO2). واثناء الحالة العجينية الرطبة، ستزيد الخمائر من المحتوى البروتيني، وستعوّض نقص بعض الحموض الامينية وتقوم بتركيب فيتامينات جديدة، اي انها تزيد من القيمة الغذائيّة. يمكننا ملاحظة مشهد سابق لتحضير الجعة، من خلال مشروب مستوطن في منطقتي النوبة والسودان، هو مشروب bouza والمحضّر اعتباراً من طحين القمح. في تحضيره، يتم تحويل ثلاث ارباع الكمية من القمح الى عجينة مع خميرة إثر تعريضها للنار الخفيفة، حيث يُسمح لتلك الفطريات متابعة عملها. من جانبه، يكون الربع المتبقي من القمح رطباً ومعرضا للهواء الجوي، قبل دقّه { خطوة تذكرنا بتحضير المولت }. لاحقاً، يجري خلط العجينة كما الربع المدقوق ويُتركان للتخمّر في وعاء مع الماء. لاجل استهلاكه، فيما لو يتم تصفية المُنتج النهائي، نحصل على مشروب شبيه بالجعة او بالبيرة، لكن سيكون لدينا غذاء عالي القيمة والذي يذكرنا بأكلة migas { عبارة عن خبز مفتت ومقلي ومغموس بالحليب .. قاموس المورد اسباني عربي }. نرى من خلال هذا المُنتج اتصال وثيق بين تحضير الخبز والجعة، هذا المشروب الذي تحوّل ليكون عنصر اساسي في غذاء مجتمعات الهلال الخصيب ومصر القديمة. حتى انه وفق بعض الباحثين يمكن القول بأنّ الخبر والجعة كانا عنصرين قد تمّ تحضيرهما في ذات المكان. في مصر القديمة، نعثر على الادلة المبكرة على تحضير الجعة في عصر ما قبل الأُسَرْ خلال الفترة { 5500 الى 3100 عام قبل الآن }، ولو أنّ كثير من الباحثين يعتبرون الهلال الخصيب الموطن الاساس للجعة في ارض الرافدين منه بالتحديد، حيث ستكون احدى ثمرات بابل منذ 8000 عام قبل الآن. فليس صدفة عثورنا على احد اقدم الادلة على تحضير الجعة في موقع Tepe Gawra بشمال العراق، والذي يعود الى 6000 عام قبل الآن. من حفريات قرب منطقة نينوى بموقع اسمه Tepe Gawra شمال العراق قبل 6000 عام،عبارة عن ختم عليه شخصان يشربان البيرة أو مشروب شبيه لقد تحولت الجعة الى مشروب شعبي في مصر القديمة والهلال الخصيب، حيث كانت تحضر ليس من الشعير فحسب، بل من انواع مختلفة من القمح كذلك. كانت تستهلكها كل الطبقات الاجتماعية، سهلة التحضير { قياسا بخمور اخرى كالنبيذ مثلا،فاشجار العنب لا تكون سهلة الزراعة والانتاج كما تكون البقول }، كما انها كانت تُعتبر بمثابة غذاء اوليّ. رواتب ومهور اعراس كانت تُدفع على شكل جعة. كما انها كانت تُستعمل كدواء أو مستحضر تجميلي لعلاج بعض الحالات كالانزعاج المعدي او السعال وغيرها. شكّلت البيرة أو الجعة العنصر الفاعل في الاحتفالات والسهرات. لقد بلغت اهميتها حدّاً نالت بموجبه تحضير مراسيم شرعنة. كان يحضر في خمّارات الجعة بشرق المتوسط الموسيقى والدعارة. حتى اننا نجد في قصيدة سومرية، الآتي: كانت الاعضاء الجنسيّة للإلهة إنانا ، إلهة الحبّ والحرب بارزة " كحلوى مثل الجعة " . لانّه حتى الآلهة لم تستطع مقاومة الجعة. فكانت نينكاسي Ninkasi هي إلهة الجعة والكحول، ايضاً نجد كمرافقين: الإله دوموزي، إله الخصب البابلي، الإله إنليل,، الإلهة المصرية Hathor أو الآلهة اليونانية Ceres و Dioniso. حتى أنّ السلف المُباشر لألوهة الثقافة الغربية التقليدية، يمكن ان يكون أكبر شارب للجعة، يناقش للآن بعض الباحثين فيما لو أنّ المقياس hin اليومي حيث (1 hin ~ 4 litros) الذي وضعه يهوه بين الصدر والظهر: كان جعة أو نبيذ، حسناً، أبعد من يهوه، لدينا كهنته { سفر العدد 28:7-10 } الذين اكتشفوا منذ زمن طويل فضيلة القربان! هل تحتاج الصورة لشرح؟فالصبيّة تشرب الجعة.لوحة من بابل القديمة 1.800 قبل الميلادمعروضة في المتحف البريطاني  أباريق جعة، عُثِرَ عليها في فلسطين وتعود لعصر الحديد أثر مصري قديم من زمن الأسرة الثامنة عشرة، يعود إلى 1.350 قبل الميلاد، حيث يشرب مهني الجعة |
|
|
|
|
|
|
08-07-2016, 09:52 AM
|
رقم الموضوع : [7] |
|
عضو ذهبي
|
البُستان التطوريّ {7}: تطور الحُمُّص والعدس  يمتلك هذا الموضوع الجديد من سلسلة " البُستان التطوريّ " شيئاً مُختلفاً عمّا حدث في المواضيع السابقة والمنشورة حتى الآن، فلأوّل مرّة سنتناول الحديث حول نوعين لا نوع واحد، بذات الوقت. وهذا يحصل لسببين: السبب الأوّل: لم تكن التغيرات التي مرّ بها كلا النوعين شديدة كالتغيرات التي رأيناها في نباتات سابقة. السبب الثاني: يشكلان نوعان مُتعارضان بشكل لبق فيما بينهما. يمكن ان نُضيف سبب ثالث: فقد حان الوقت للتكلُّم عن القطّانيّات { الحبوب التي تُطبخ كالعدس والحمّص والفول واللوبياء .. قاموس المورد اسباني عربي }! العدس والحمّص، اضافة للفاصولياء، الصويا، البازلاء، الفول وغيرها: تشكّل نباتات تنتمي الى عائلة القرنيّات Leguminosas، وهي مجموعة ناجحة تشغل الترتيب الثالث في عدد انواعها بين كاسيات البذور angiospermas { النباتات الزهريّة }، وتأتي بالضبط خلف السحلبيات orquídeas { فصيلة نباتيّة من وحيدات الفلقة منبتها الاقاليم الحارة .. قاموس المورد اسباني عربي } و المُركّبات compuestas { فصيلة نباتية من ذوات الفلقتين وحيدة القُعالة سفليّة الاسدية تُعتبر من اكبر الفصائل النباتية .. قاموس المورد اسباني عربي }. ليس صدفة ان نعثر على 20000 نوع تقريباً حتى الآن ضمن 700 جنس. من الامور المثيرة أنّ قطانياتنا المُدجّنة قد سارت في خطّ شبيه بخطّ بقولياتنا { قمح، شعير، ذرة وأرز } خلال عملية تدجينها، ما يجعلها مفيدة لنا انطلاقاً من وجهة نظر اصول نباتيّة وتطوريّة.  الى اليسار حفنة من العدس،الى اليمين حفنة من الحمّص تمتلك مجموعة القرنيات اسماء كثيرة، فهي تسمى قطانيات واصلها اللاتيني «lĕgūmen» عبارة عن مصطلح معناه " الذي لا يتغيّر بمرور الزمن ". ايضا تكون معروفة تحت اسم fabáceas القادم من المصطلح اللاتيني «faba»، وهي كلمة ما تزال مُستخدمة لليوم في شمال الجزيرة الايبرية للدلالة على الفاصولياء، مع انّه بالاصل قد أُطلِقَتْ كلمة «faba» الرومانيّة على الفول. كما تسمى باسماء اخرى، احدها يأتي من المُصطلح اللاتيني «papilio» اي { الفراشة } وذلك بفضل شكل ازهارها الجميلة. تتكوّن تلك الازهار من 5 بتلات pétalos متموضعه بصيغة خاصة، اثنتان منها ملتحمتان جزئياً، حيث تغطيان اعضاء التكاثر بالزهرة وتشكلان ما يشبه القاعده quilla، بتلتان اخريتان حرّتان محيطتان بتلك القاعده هما جناحان alas، وبتلة اخيرة ذات حجم كبير تموج خلف البتلات الاخرى وتسمى الراية estandarte. بالتفافها حول القاعده، يكون لدينا اعضاء التذكير " الأسدية " androceo والتي تُحيط بدورها مجموعة اعضاء التأنيث في الزهرة، يتكوّن الجهاز الذكري من عشرةgineceo ± أسدية، والتي يمكن ان تكون كلها حرّة، كلها مُلتحمة فيما بينها أو كلها مُلتحمة ما عدا واحدة منها.  جدول اساسي لزهرة القرنيّات.كمثال لدينا نوع البازلّاء Pisum sativum L مقدمات {1} العدس يُعرف العدس المُدجّن الذي نتناوله في العالم الآكاديمي تحت اسم: Lens culinaris ssp culinaris Meiken انه نبات صغير ذو مظهر شجيريّ، يبلغ طول ساقه من 20 الى 30 سنتمتر. يمتلك دورة حياة سنوية، يمكن ان يشهد نموّ سريع، حيث يبلغ سنّ النضوج خلال فترة تمتد من 75 الى 100 يوم فقط. تُعطي ازهاره البيضاء الصغيرة بعروقها الزرقاء الاصل لظهور بذرة أو بذرتين رقيقتين بشكل عدسة. تكون تلك البذور غنيّة بالبروتين، الاملاح المعدنية { صوديوم، بوتاسيوم، حديد وزنك } وفيتامينات { بترتيب اهميتها: ب، ث، آ، ي و ك } وهي غذاء لا يُستغنى عنه. اضافة لانّه وبفضل احتوائها المُرتفع من الاحماض الامينية كالليسين lisina والتربتوفان triptófano، اذا تمّ جمعها بالقمح او بالأرز فإنها تُكمل احتياجاتنا الى الحموض الامينية الاساسيّة. { في منطقة شرق المتوسط يوجد أكلتان شهيرتان تُصنعان من البرغل والعدس، والارز والعدس، واسمهما الشعبي " مُجدّرة " وهما اهمّ تعبير عن صحيّة غذاء اهالي تلك المنطقة .. ودون علمهم بهذا طبعاً .. فينيق }. يوجد اهتمام بالغ بزراعة العدس. فهو يُزرع بأكثر من 40 بلد، وقد بلغ الانتاج العالمي منه 3.8 مليون طن سنويا بين العامين 2004 و2006 { بحسب معطيات منظمة الفاو 2008 } وبمساحة مزروعه بلغت 4 ملايين هكتار. أهم ثلاث مُنتجين للعدس: الهند، كندا وتركيا، بينما اوروبا كاملة لا تُنتج بالكاد 1% من الانتاج العالمي من العدس.  نموذج من العدس Lens culinaris culinaris Medik يشكّل العدس جنس نباتيّ قليل التنوّع حيث يتواجد، يكون مزوّداً في 7 ازواج من الكروموزومات (2n = 14، يتوزّع حول البحر المتوسط. الانواع البريّة التي تنمو في شبه الجزيرة الايبرية، هي: Lens nigricans { متوفرة نسبياً } و Lens lamottei { موجودة في قطاع المتوسط }، بينما في جزيرة مايوركا { جزر الباليار } نجد نوع Lens ervoides. Lens culinaris، النوع المزروع المُتاصِّل في المتوسط الشرقيّ.  تنوعات الجنس البرّي Lens وتوزعه الجغرافي في العالم هنا يحضر سؤال حيويّ انطلاقاً من وجهة نظر تاريخيّة: ما هو السلف البريّ لعدسنا المزروع؟ كان اول شخص انطلق في البحث عن إجابة، هو عالمة النبات الروسيّة Elena Ivanovna Barulina، طالبة وزميلة ولاحقاً زوجة عالم النبات والوراثة الروسي Nikolái Ivánovich Vavílov. كانت تتركّز ابحاث العالم Nikolái Ivánovich Vavílov حول معرفة اصل وتطور نباتاتنا المزروعه. تأسيساً على ميزات تشريحية، اقترحت العالمة Elena في العام 1930 بأنّ السلف البرّي للعدس Lens culinaris، هو المعروف تحت اسم Lens orientalis { اليوم معروف باسم Lens culinaris ssp. Orientalis } ذو اصل شرق متوسطيّ. وللآن كثير من الباحثين متفقين مع هذا الطرح. فلقد أثبتت دراسات البروفيسور Ladizinsky { سنتكلم عنه عند حديثنا عن الحمّص } بأنّه تمّ تهجين نوع L. culinaris مع نوع L. orientalis دون وجود اي مشكلة { بعكس ما حدث مع كل انواع الجنس Lens }. حديثاً، تمّ تأكيد هذا الطرح وبشكل متكرر بواسطة علم الاحياء الجزيئيّ biología molecular. كيف تكون تلك النبتة؟ بخلاف النباتات الاخرى التي درسناها بهذه السلسلة، يكون نوع Lens culinaris ssp. Orientalis متطابق تقريباً مع نسخته المزروعة. يكون اقلّ حجماً فقط بعدد الوريقات في الورقة، في مساحة تلك الوريقات تغطي " شعيرات " اوراقها وسوقها، نادراً ما تُعطي ازهار بيضاء، تكون بذوها صغيرة جداً { حيث تكون الميزة الوحيدة التي تفرقها عن نسختها المزروعه في الطبقات الاثرية } ..الخ.  مثال دقيق لنوع العدس Lens culinaris ssp. Orientalis {2} الحُمُّص يكون اسمه العلميّ Cicer arietinum L، عبارة عن نمط شجيري لا يتجاوز ارتفاع نبتته النصف متر, وتكون ثماره عبارة عن حبّات او بذور، تُحاط كلّ حبّة واحدة بسنفة { السِنفة هي وعاء حبّ البقل كالفول والعدس والحمص وغيرهم .. قاموس المورد اسباني عربي }. تمتلك تلك البذور قيمة غذائيّة عالية، انها غنيّة بالبروتين، الالياف، املاح معدنية { نحاس، منغنيز، حديد وزنك } والفيتامينات { بترتيب الاهمية: ب، ث، آ، ي وك }. وكالعدس، انها مًكمّل ممتاز للبقوليات. يشكّل Cicer arietinum النوع الوحيد المزروع من جنسه. عبارة عن نبات يُزرع في 49 بلد. يبلغ انتاجه العالمي حوالي 8.6 مليون طنّ سنوياً { وفق معطيات منظمة الفاو 2006 } وتتم زراعته على مساحة قدرها 11.2 مليون هكتار. اهم البلدان المنتجة له، هي: الهند، الباكستان وتركيا، فيما تحتل اسبانيا الترتيب الحادي عشر بين الدول المُنتجة له. بكل الاحوال، بالكاد يشكّل الانتاج الاوروبي له ما نسبته 1% من الانتاج العالمي.  نموذج من الحُمُّص Cicer arietinum L ينضوي تحت الجنس Cicer 44 نوع { 9 ذو دورة حياة سنوية و35 ذات دورة حياة دائمة } وتنتشر في جنوب غرب آسيا. يوجد في اسبانيا نوع بري واحد فقط من هذا الجنس ويكون مستوطناً. النوع هو Cicer canariense A. Santos & G. P. Lewis، عبارة عن عشبة دائمة تعيش في اماكن رطبة على ارتفاعات 420 – 1535 متر عن سطح البحر. انه نوع مستوطن حصرياً في جزيرتي لاس بالماس وتينيرفي { ضمن جزر الكانارياس }، مصنّف بكونه " سريع العطب " وفق كاتالوك الانواع المهددة بالانقراض في جزر كانارياس، وبحسب القائمة الحمراء الصادرة العام 2008 حول الازهار الوعائيّة الاسبانيّة.  نموذج من الحمّص البرّي المُستوطن في كانارياس Cicer canariense الموجود حصرياً في جزيرتي Palma و Tenerife سنتبع ذات الخطوات مع العدس. ما هو أصل الحمّص؟ يأتينا أول الاجوبة من طرف البروفيسور Gideon Ladizinsky من معهد علم وراثة النباتات في الجامعه العبرية في القدس المحتلة. حيث تركّزت ابحاثه حول التغيرات التطورية التي اصابت النبتات خلال عملية تدجينها، وليس مصادفة ان يكون مؤلّف كتاب Plant Evolution Under Domestication المنشور في العام 1998. بحسب البروفيسور ودراسات لاحقة تابعت خطواته، يكون السلف البري للحمّص المزروع، عبارة عن نوع نادر من الحمص البري والمُكتشف من قبل البروفيسور ذاته والموجود في تركيا حصرياً. اسمه العلمي Cicer reticulatum Ladiz. وذات الامر يحصل مع العدس، في الشكل العام تكون شبيهة كثيرة مع نسختها المزروعه. ينمو النوع Cicer reticulatum في اراضي كلسيّة على ارتفاع 600-1500 متر عن سطح البحر، مشكلاً لجماعات منتشرة مختلطة مع بقوليات اخرى باراضي جافة تستقبل سنويا ما معدله 350 – 830 مليمتر من الامطار. تمّ اثبات أنّ النوعين C. reticulatum و C. arietinum خصيبين فيما بينهما، ما يعني، أننا فيما لو نقم بتهجينهما سنحصل على متحدّر خصب وقابل للحياة، في الواقع يشكّل C. reticulatum النوع الوحيد الذي يمكن تهجين حمصنا المزروع معه بنجاح. مع ذلك، لا يحدث هذا التهجين بصورة طبيعية اطلاقاً، والصيغ الهجينية { حتى متى كانت الصيغ المزروعه والبريّة قريبة جداّ مكانياً } لا يُعثر عليها. هذا يحصل لانّ كلا النوعين C. reticulatum كما C. arietinum، يشكلان نباتان ذوي تلقيح ذاتيّ بارز، اي انهما يخصبّان ذاتهما ويتم نشر غبار طلعهما عبر الرياح او الحشرات لاجل القاح ازهار اخرى. لهذا السبب يعتبر دارسي تلك النباتات، كما يمكننا رؤية هذا في المراجع، الحمص المزروع و " الحمص الشبكيّ " كنوعين مختلفين.  تفاصيل زهرة لنوع Cicer reticulatum Ladiz ، انها صورة من ضمن قليل من الصور التي امكنني العثور عليها لهذا النبات سفرة مُقتضبة عبر الزمن متى بدأ تدجينه؟ كيف حدث ذلك؟ ليسا سؤالين سهلين. اول ما يمكن الاجابة عنه، هو أنّ التدجين قد مرّ بخطوات مختلفة بكلا النوعين. في جزء تبعاً لعدم انتظام توزعه الجغرافي، فمساحة توزع " العدس الشرقي " كبيرة، بينما يعيش " الحمص الشبكيّ " في تركيا ومناطق قليلة أخرى. كما هو معتاد في هذه السلسلة، من جديد تقودنا آلة الزمن نحو منطقة الهلال الخصيب Creciente Fértil، هذا المكان الممتليء بالحياة منذ نهاية آخر الحقب الجليدية { منذ 11000 عام } من شرق المتوسط وصولاً الى مصر القديمة. ارض سابحة في النيل، نهر الاردن، دجلة والفرات، حيث عاشت نباتات سنوية وانتشرت بشكل متكرر. يُعتبر الهلال الخصيب مهد الحضارات في العالم القديم، والمكان الذي وُلِدَتْ فيه الزراعه. ونحن اليوم نتابع اسهامنا بتلك المغامرة.  خارطة الهلال الخصيب منذ ما قبل 13.000-11.000 عام تظهر اوائل المؤشرات على استخدام العدس من خلال بذور متفحمة، جنبا إلى جنب مع حبيبات القمح والشعير البري، في طبقات العصر النيوليتي قبل السيراميكي في تل المرابط وتل ابو الحريري { الجمهورية السورية } وموقع Netiv Hagdud { فلسطين المحتلة } والتي تعود للفترة 9500-7200 عام قبل الآن. تمّ العثور على اكتشافات شبيهة في كهف Franchthi { اليونان } وفي Grotta dell’Uzzo { في صقليّة } حيث ينتمي العدس المعثور عليه بالتأكيد الى النوع المحليّ الوافر Lens nigricans. اعتباراً من 7200 قبل الميلاد، يُصبح مألوفاً العثور على بذور متفحمة من العدس في اماكن عديدة من الهلال الخصيب. وبناء على صغر حجم تلك البذور { 2.5-3 مليمتر }، يكون مستحيلاً تحديد فيما لو كانت بريّة أو انها نتاج بدء زراعتها. فقط في وقت متأخّر، وفي طبقات العصر النيوليتي ما قبل السيراميكي في موقع يفتاح { شمال فلسطين المحتلة، يعود الى تاريخ 6800 قبل الميلاد } يبدأ العثور على بذور عدس " متلوثة " بنوع نباتيّ هو Galium tricornutum يصاحب بيومنا هذا النوع المزروع من العدس، ما يعني ان تلك الشعوب قد بدأت على الارجح بزراعة العدس منذ اكثر من 8000 عام، إثر ذلك التاريخ وبمرور 1000 عام تبدا زراعة القمح والشعير. ربما يكون صائباً اعتبار بداياته الألف السادس قبل الميلاد، حيث يتم العثور في طبقات موقع Tepe Sabz { إيران 5500-5000 قبل الميلاد } على بذور عدس بقطر قدره 4.2 مليمتر. وهو حجم يراه علماء النبات أكبر بكثير من النسخة البريّة، ما يعني ان تلك الثمار قد جرى تحصيلها من الانتقاء قبل ذات التاريخ بزمن غير قليل. بذات الحقبة نعثر على بذور في موقع Khirokitia { في قبرص }، وهو حضور يشكّل نتيجة لانتشار باكر لزراعه هذا النبات وصولا لاوروبا. بطول الالفية الثانية قبل الميلاد، امتدت زراعته الى افريقيا وآسيا. بمرور الزمن وتطور الحضارة، تحوّل العدس الى غذاء اساسي للطبقات الاكثر فقراً في اليونان القديمة { 455-387 قبل الميلاد }، بينما اكتسب في روما صفة " غذاء طقسيّ " اثناء حفلات الدفن.  الاماكن الرئيسية لانتشار زراعة العدس، فترة ما قبل الميلاد أما بالنسبة للحمّص، فربما يعود تاريخ بدء زراعته الى 10000 عام قبل الآن. تُعتبر البذور الاكثر قدماً من الحمص المؤرّخة بالالف الثامن قبل الميلاد، وتنتمي تلك البذور الى طبقات موقع تل ابو الحريري وتل الكرخ { في الجمهورية العربية السورية حالياً }. حيث تُظهر تلك البذور شكلا متوسطاً بين النموذج البري والنموذج المزروع. نقطة اخرى بصالح تطبيقات زراعية خضعت لها تلك البذور، حيث انها رُصدت على بعد 260 كيلومتر عن نطاق توزعها الطبيعي. وكدليل مضاد، يؤكد بعض الباحثين بأنه ربما تكون الجماعات الحديثة بقايا توزّع طبيعي وانه كان اكثر انتشاراً في الحقب القديمة. يُلاحظ الانتشار الواضح للحمّص بحدود الالف السابع – الألف السادس قبل الميلاد، حيث يظهر في طبقات العصر النيوليتي ما قبل السيراميكي بموقعي Jericó و Ramad مكان بعيد عن مركز اصله حيث يتم الحديث عن زراعته. ايضاً تمّ العثور على بذور محمصّة من نوع C. reticulatum في طبقات موقع تركي اسمه Çayönu { 7500-6800 عام قبل الميلاد } وموقع تركي آخر اسمه: Asikli Höyük { 6900-6500 عام قبل الميلاد }. يبدأ توفّر هذا النبات بكثرة في طبقات الهلال الخصيب، ابتداءاً من العصر البرونزي الباكر { 3300-2200 عام قبل الميلاد } في فلسطين والاردن. تشكّل هذه بالنسبة لعلماء الآثار شاهد واضح على زراعة الحمّص، حيث تختلف بذورها عن بذور النوع الوحيد من الحمّص البري الموجود في ذات المنطقة وهو Cicer judaicum. بعد زراعته بزمن طويل، انتشر حتى بلغ جزيرة كريت ومصر، وفي عصر الحديد { 1000 قبل الميلاد } امتدت زراعته لتصل الى اثيوبيا وغرب آسيا.  الاماكن الرئيسية لزراعة وتوزع الحمّصاختصار بفترة قبل الميلاد تطور العدس والحمّص وفق عملية التدجين العدس من جديد فيما لو يكن من الصعب معرفة زمن ذروة استخدام نبات ما، أو متى بدأت زراعته، فإنّ الاكثر تعقيداً هو معرفة اسباب اختيار هذه الانواع للزراعة. ودون الذهاب بعيداً، فكثير من البقول مثل Lens culinaris ssp. Orientalis تُبرز مساويء واضحة، منها: أولاً: قليلة الانتاج. قام بعض الباحثين بحساب انتاج النبتة، ليصل الى: 1-45 بذرة وبمتوسط قدره 10 بذور للنبتة الواحدة. ما يعني انه لجني كيلو واحد من العدس، يجب قطاف 10000 شتلة تقريباً. وبناء على هذا الجهد المبذول بقطافه، يجب ان يشكّل العدس جزء صغير من النظام الغذائي لمجتمعات العصر النيوليتي ما قبل الزراعي، ولهذا السبب ربما، كان هذا النوع غير لافت للانتباه في الزراعة. ثانياً: من الصعب تحقيق انتاجيّة متواصلة. حيث تعاني بذور النوع L. culinaris ssp. Orientalis من حالة " نوام "، وهي عبارة عن حقبة سُبات تمنع موت البذرة عبر تفادي انتاشها بفترة جفاف. يتحقق " النوام " بفضل غلاف كتيم يحمي البذرة. فيما لو يكن الغلاف ملغي، كذلك سيكون " النوام " ملغي. مع هذا، رغم فعالية هذا الغلاف، نجد ان 10 الى 15% فقط من البذور تصل لمرحلة الانتاش في العام التالي. وهذا يشكّل دمار من وجهة نظر زراعيّة، حيث يتحول كل الانتاج الى موسم الزراعة في العام التالي .. حيث سنحقق ذات الانتاج.  حقل مزروع بالعدس.فاليوم لا تُواجهنا مشاكل كثيرة.منطقة بريدا، سورية، الفترة 1998/1999 بفضل اعمال الدكتور Ladizinsky نعرف اليوم بلزوم جين واحد فقط لتحديد " النوام "، بالتالي تحقيق طفرات حادثية بهذا الجين، سيكون بامكانها الغاءه. يتفق بعض الباحثين بكفاية حدوث هذا تحت ظروف مائية { من 4 الى 7 اعوام ماطرة دون جفاف }، كي تسود النبتات " الغير نائمة " في الجماعة بفضل نموها السريع وايقاع تضاعفها العالي. سيطرة ستستمر حتى حقبة الجفاف التالية. يعتبر عالم النبات Daniel Zohary بانه بمجرد تحقيق تلك الظاهرة، بدأت زراعة العدس وبسرعة، حيث سادت " الغير نائمة " في عمليات الزراعة. من جانبه Ladizinsky يبرهن بأنّ هذا لا يحلّ مشكلة انتقاء ذلك النوع والحفاظ اللاحق على زراعته المعطي وفق مردوده القليل. لهذا يؤكد بأنّ الغاء " النوم " قد حدث قبل تدجين العدس، بحسب زراعتها، بحقبة تمتد من 4 الى 6 اعوام دون جفاف، سيكون كافياً لتبلغ العدس " الغير نائمة " اكثر من 50% من العدس الكليّ. ليس فقط هذا، بل بالاخذ بعين الاعتبار للقدرة الضئيلة على نشر البذور، لن تشكّل نبتاته الكثيفة سهولة في الجني فقط، بل ستشكل حافز في الاهتمام المستقبلي بزراعته. تغيّر جوهري آخر في البقوليات والقطّانيات المزروعه، هو تثبيت البذور الناضجة على النبات البالغ. فلقد رأينا ان هذا قد تحقّق في البقول من خلال السنابل غير الهشّة، والتي تحفظ البذور حتى وقت قطافها بدل ان تكون منثورة بواسطة الرياح أو الحيوانات. حصل شيء مشابه في القطّانيات. حيث تنفتح السنفات الناضجة ذاتيا محررة محتواها، انها منفتحة dehiscentes. بينما يكون الامر معاكس في بقولياتنا المزروعه، فالبذرة محتجزة في سنفة لا تنفتح عند نضجها، انها غير منفتحة indehiscentes. في العدس، يكون منفتحا dehiscencia ميزة مسيطرة امام الميزة غير المنفتحة indehiscencia، ورغم ذلك فإنّ الميزة الاخيرة تلك سائدة في عدسنا المزروع. ويمكن تفسير هذا بوصفه نتيجة انتقاء مفروض عبر التدجين. وفّرت ابحاث Ladizinsky في العام 1979 الجدول التالي، حيث نجد بعض الميزات المختلفة الاكثر اثارة بين النوعين البري والمزروع، على الصعيد الجيني الوراثي.  جدول يوضح توريث الميزات التشكلية في نوع العدس L. culinaris. Scp بقع في غلاف البذرة، Gh نمط النموّ، Gs لون epicótilo ، Pi سنفة منفتحه بحسب Ladizinsky في العام 1979 مع هذا، تبين دراسات حديثة بأنّ نتائج الجدول السابق تُعاني من نقص. تمّ التحقّق من الفروقات { بين العدس المزروع والعدس " الشرقي " } في 9 ميزات { ارتفاع العقدة الاولى nodo، عدد التفرعات حتى العقدة الاولى، العدد الاجمالي للتفرعات، ارتفاع النبات، زمن الازهرار، منفتحة، عدد البذور، وزن البذور وقطر البذرة } والمتوجبة على 23 QTL «Quantitative Trait Loci» التي كانت متبدلة. ما يعني، انهم قد اكتشفوا بأنّه على الاقلّ 23 مقطع من الحمض النووي DNA تكون مرتبطة بتحضير تلك الميزات التسع، قد خضعت للتعديل خلال آلاف الاعوام الاخيرة من التدجين. نجد بالتالي انه لاجل كل فارق بين العدس " الشرقي " و العدس " المزروع "، يوجد { بالتأكيد } أكثر من جين واحد متعدّل. كمثال، راينا في الجدول السابق بأنّه اعطي تفريق ميزة الانفتاح dehiscente بنسبة 1:3 مع ذلك بحسب تلك الدراسة لا تكون النسب بكل هذه الدقة، بل تتغير بصيغة تبرز حضور 3 QTL { مثل الصويا نوع Glycine max } متدخلة حول الميزة، والتي يُفسر علماء الوراثة تأثيرها عبر جين رئيسي وجينان ذوي فعل ثانوي.  بعض انواع العدس سؤال هام آخر يحضر هنا.. لماذا هناك بذور ذات لون اصفر .. ضارب الى الرمادي؟ من حيث المبدأ، هذه ميزة غير هامة كثيراً، في الواقع لاجل البرهان على هذه النقطة، سنفترض بان تلك الميزة غير هامة زراعيا ولا غذائياً. في حال كان الامر هكذا، لماذا يتم انتقاء تلك الميزة؟ لماذا تبرزها النباتات المدجنة؟ يكون هذا مناطاً ب " جينات لا تشكّل ذاتيات حرّة ". تشكّل الجينات تعاقبات من الحمض النووي DNA مندمجة في الكروموزومات { الصبغيات }. عند العثور على جينين بذات الكروموزوم، يمكن ان يوجدا فيزيائيا الواحد بجانب الآخر. لهذا تسمى بجينات مرتبطة، وحيث انهما سوياً، يورثان سوياً. لكن في جينوماتنا لا نمتلك زوج من الجينات فقط، بل اكثر بكثير منهما. عند وجود جينين في كرومزومين مختلفين، فاننا نتحدث عن جينومات غير مرتبطة حيث انهما منفصلان، يورثان منفصلان. حيث ان الميزات الشهيرة التي درسها ماندل في البازلاء تتقاسم الخيار الاخير ذاك. هذا يفسّر لنا سبب كون ميزات محددة لا تمتلك اهمية زراعية، مثل: حضور أو غياب الشعرانيّة في السنفات { السنفة هي الغلاف المحيط بحبوب البقليات، كقرن الفول والبازلاء والحمص والعدس .. الخ }، لون الاوراق او عدد الوريقات بكل ورقة، تكون منتقاة. لن تكون تلك الميزات هامة، لكنها تصبح مترابطة ببعضها، وترتبط بشيء هام كالتنظيم الجيني لحجم النبات. بذات الصيغة، يرتبط لون سطح البذرة والوريقات بالاوراق بالجينات التي تحدد الإزهرار المبكر.  فرق بين الجينات المستقلة غير المرتبطةوجينات مرتبطة التي تورث سويا الحمُّص من جديد لدينا اشياء يجب قولها بخصوص الحمّص نوع Cicer arietinum. مرّت كل النباتات خلال عملية تدجينها بظاهرة تسمى " عنق الزجاجة "، حيث يحصل خسارة كبيرة في التنوع الجيني بسبب اعطاء الاصل لاوائل النباتات المدجنة من خلال افراد قلائل باماكن محددة حصرياً. يعني حضور افراد قلائل امتلاك تمثيل صغير جداً للتنوع الجيني الاجمالي للنوع الحيّ. هذا النقص بالتنوع الجيني { وبجينات مفيدة بشكل فعّال } يمكن تخفيفه بجانب مع هجائن بين انواع مزروعة ذات اصل مختلف و/ أو مع هجائن مع انواع بريّة. كان " عنق الزجاجة " أكثر وضوحاً في الحمّص. ففي المقام الاول، لان الجماعات الاصلية للسلف البري Cicer reticulatum قد كانت ضحايا لظاهرة " عنق الزجاجة ". في المقام الثاني، بينما قد تنعّم العدس وبقوليات اخرى { كالبازلاء } بمواطن تدجين عديدة، فإنّ الحمّص قد تمّ تدجينه باماكن قليلة في تركيا. في الواقع، يمتلك الحمّص المزروع تنوع جيني اقلّ من نظيره البري.  توزع جغرافي لانواع اسلاف لانواعنا المزروعه.في اليسار: الحمّص.في الوسط: الشعير.النجيلات البرية،القمح القاسي.في اليمين: العدسوالبازلاء لماذا توجّب اختيار نوع الحمّص؟ يمتلك نوع Cicer reticulatum فائدة لا تمتلكها البقوليات الاخرى التي عاشت في الهلال الخصيب، فاضافة الى حجمه الاكبر، فهو يمتلك سنفات منغلقة indehiscentes تقوم بحفظ البذور حتى بعد بلوغها بزمن طويل. بحسب Ladizinsky، يحوله هذا الى مصدر حيوي للغذاء، وبناء عليه تصبح جاذبية زراعته اكبر. نقطة اخرى يقدمها فريق بحثي بقيادة Shahal Abbo من معهد علوم النبات والوراثة الزراعية بالجامعه العبرية في القدس المحتلة، حيث اولى اهتمام خاص حول تاريخ الحمّص، وبحسب هذا الفريق: توجّب اختيار الحمّص بسبب قيمته الغذائية العالية. فالحمّص مصدر لحمضين امينيين اساسيين { triptófano و fenilalanina } يساهمان برفع مستوى الرضى والشبع، انها هدية بيولوجية تتمثل بنوال غذاء اساسي لاجل عمل جيد للدماغ. في الواقع، يكون حمصنا المزروع غني بشكل مضاعف بالحمض triptófano قياسا بالنوع البري. أليس دليلاً اضافيا على اهمية الفعل الانتقائي والغير واعي للقيم العليا للحموض الامينية الاساسية؟ بالتأكيد.  جدول يبين القيمة الغذائية اليومية من الحمص بالغرام للحصول على اللازم يوميا من الحمض triptófano وفق منظمة الفاو يبرز " عنق الزجاجة " التالي الذي تعرّض له الحمّص، من خلال: رهان خطر قد غيّر دورة حياته. تمتلك كثير من نباتات المتوسط، وعلى وجه الخصوص نباتات الهلال الخصيب: دورة حياة يحصل فيها الانتاش بفصل الخريف وتنمو حتى نهايات الشتاء { حيث تستقبل 80% من الهطولات المطرية السنوية }، ويحصل الازهرار باوائل فصل الربيع وتصل الى البلوغ قبل وصول الصيف. يشكّل الحمّص المزروع استثناء بهذه القاعده، فهو يُزرع بنهاية الحقبة الماطرة وينمو خلال قيظ الصيف الشديد. لا يكون هذا مجانيّ. ففي بعض مناطق فلسطين المحتلة للآن يُزرع الحمص في فصل الخريف، حيث يبلغ الانتاج 3 طن للهكتار، وهذا اكثر بكثير من 700 كغ للهكتار للنوع المزروع في الربيع. حسناً، لماذا كل تلك التضحية بهذه الكمية الزائدة من الانتاج اذاً؟ لماذا يتم اتباع هذه الاستراتيجية التنافسية؟ يكون السبب جوهري، فهناك نوع من الآفات الشديدة الخطر، نوع من الفطور المسماة Didymella rabiei. حيث تهاجم القسم الهوائي من النبات { بذور، سنفات، ازهار، اوراق وساق } مسببة مرض يسمى " داء كلب الحمّص "، حيث تتضمن عوارضه ظهور بُقع ومساحات منخورة. تكون مؤثراته فظيعة الى درجة تخريب كامل المزروعات منه. ولحسن الحظ ان هذه الآفات تكون ضعيفة ولديها معيقات للتاثير في فصل الربيع، وهذا بفضل الرطوبة البيئية المسيطرة.  تاثيرات الفطر Didymella rabiei على الحمّص  بعض أنواع الحمُّص  حقل حمُّص في استراليا  آثار الطفرات والتطوُّر. في الأعلى وإلى اليسار زهرة بيضاء متطفِّرة وإلى اليمين زهرة بريّة. في الأسفل وإلى اليسار بذور بريّة، في الوسط بذور متطفِّرة وفي اليمين بذور مزروعة يتبع |
|
|
|
|
|
|
08-11-2016, 05:56 PM
|
رقم الموضوع : [8] |
|
عضو ذهبي
|
البُستان التطوريّ {8}: تطور دوّار الشمس  يكون اسمه العلميّ الشهير هو Helianthus annuus L ويشكّل اسم مُعتمَدْ بشكل حرفيّ، حيث ان الاسم Helianthus مُشتقّ من مصطلحات يونانيّة helios, sol و anth(us) flor وهي اشارة مباشرة لسلوك هذا النبات وفق التوجُّه الشمسيّ heliotropismo أي توجّه الازهار والاوراق نحو الشمس. أما annuus فيعني باللاتينيّة anual وسنويّ بالعربيّة ويُشير لدورة النموّ والتكاثر لهذا النبات حيث انه بتلك الحقبة الزمنية يحصل الالقاح، النمو، الازهرار، انتاج البذور والموت. فهذا النبات " الزهور الشمسيّة " ينضوي الى جانب نبات الخسّ، الأرضي شوكي، البابونج، اضراس الاسد وغيرها بمجموعة ضخمة من النباتات المعروفة باسم الفصيلة النجميّة Asteráceas ، تكون هذه الفصيلة، دون شكّ، الأكثر نجاحاً في مملكة النباتات، حيث تسجّل رقم قياسيّ بالتنوع البيولوجيّ كونها تضمّ أكثر من 20000 نوع موصوف بيومنا هذا ضمن أكثر من 1000 جنس. مع ذلكوبخلاف ما يقوله احساسنا العام، فلا تكون هذه " الورود " الجميلة لهذه المجموعة من النباتات كما تظهر.  إلى اليسار: زهرة الخسّ Lactuca sativa ، إلى اليمين: زهرة الأرضي شوكيCynara scolymus يُشتقّ مُصطلح الفصيلة النجميّة Asterácea من الكلمة اللاتينيّة aster, estrella ويكون هذا الاسم شاعريّ { بالعربيّة: نجمة }، وفي الواقع نحن لسنا امام زهور رومانسيّة تصادفنا بلحظات شاعريّة ما، انها ليست ازهار وأقوله بشكل جدّي؟؟؟!!!! في الواقع، هي تشكّل ازهرار: مجموعة من الازهار التي تشكّل باقة، ولكن في حالتنا هذه لدينا باقة مُندمجة بشكل هائل فيما بينها. وفق هذه الصيغة، فيما لو يُطلَقْ اسم سنبلة او سنابل على ازهرار القمح أو الشعير، فإنّ ازهرار النوع margaritas المنتمي للفصيلة النجمية يسمى: فصول أو فصوص capítulos. حيث حين تكون الازهار منفردة، فستكون عبارة عن كائنات صغيرة جداً جداً. كذلك يمتلك هذا الكائن الدقيق بتلات، أسدية والمياسم كأيّ زهرة أخرى، لكن بدرجة متقزمة فقط. فيما لو نقتطع زهرة ونعبر مركزها سنرى ما افترضنا أنّه كان " مركز الزهرة " الذي يشكّل قاعدة واسعة تسمى التخت receptáculo وعلى هذا التخت تجلس الازهار الدقيقة وتُعتبر كل زهرة منها عبارة عن حامل لبذرة وحيدة. هي عبارة عن ازهار تمتلك 5 بتلات ملتحمة مشكِّلة انبوب. كذلك تمتلك 5 أسدية خارجة من الانبوب هذا، كذلك تلتحم تلك الأسدية فيما بينها لتشكّل إنبوب. هكذا نجد أنّ البتلات تشكّل انبوب وضمنه تشكّل الاسدية انبوب آخر. يظهر اعتباراً من الانبوب المتشكل من الاسدية الميسم وهو العضو المُستقبِل لحبوب الطلع في الزهرة. بالاضافة لاننا نعثر بتلك النباتات على نوعين اساسيين من الازهار، تسمى احدهما lígula وتقلّد بتلة، يسمى النوع الآخر من الازهار flósculo ويقلّد السداة، لكن هل ما تزال تُعتبر ازهار كاملة ومشهدها عبارة عن تقليد محض؟!  نهج ازهرار inflorescencia مركبات الفصيلة النجميّة Asteraceae باعتبارها نباتات تؤسس استراتيجيتها على استعمال الحشرات بنقل حبوب الطلع والتلقيح،ومع درجة الاندماج بينها هذه،يتم السماح لكثير من ازهارها ان تكون محطّ زيارة بذات الوقت من ذات الحشرة. كذلك يمكن القول بناء على قاعدة نوعي الازهار المذكورين اعلاه lígulas و flósculos بأنّه يوجد ثلاث انواع رئيسيّة من الازهرار في عالم الفصيلة النجميّة، asteráceas ، ويمكن رؤية تلك الانواع الرئيسيّة في الصورة التالية، حيث يكون لدينا ثلاث أنواع تعيش بشبه الجزيرة الايبيريّة من النباتات، هكذا نجد: يمين الصورة،نوع Chrysanthemum coronarium ويمتلك lígulas باطرافه و flósculos في مركزه،انها مجموعة ازهار، ازهرار وتقليد زهرة. في حالة نبات الخسّ الذي نزرعه بحقلنا، كذلك بحالة نبات ضرس الاسد وانواع أخرى، مثل نوع Sonchus tenerrimus في يسار الصورة، نجد فصوص متكونة فقط من lígulas بالنهاية يوجد غيرها، تستقبل اسماء متعددة من قبل بعض اجدادنا، حيث تحتوي فقط على flósculos، هذا هو حال الأرضي شوكي المزروع أو نوع Senecio vulgaris وسط الصورة أدناه.  بعض ممثلي الفصيلة النجميّة في شبه الجزيرة الايبيريّة، من اليسار الى اليمين Sonchus tenerrimus, Senecio vulgaris, Chrysanthemum coronarium يبرز ملمح آخر مميّز للفصيلة النجميّة هو ثمرتها. حيث يسمى الفقيرة aquenio ويتكوّن من " جدار مقاوم " يحيط بالبذرة. لقد تشكّل هذا الجدار في الواقعاعتباراً من التحام كل طبقات الثمرة بصورة غطاء واقٍ. لتكوين فكرةK تشكّل القشرة الرقيقة والقاسية لدوّار الشمس ما يعادل قشرة ومحتوى التفّاحة. ميزة اخرى لثمار الفقيرة، هوانها لا تنفتح { بعكس غمد البقوليات } إلّا لحظة الإنتاش. وكذلك تتضمّن الفقيرات بذرة وحيدة واحدة فقط. عندما نتحدث عن الفقيرات، فنحن نُشير حصرياً لبذور دوّار الشمس. فبمجرد انخلاع القشرة تتحرّر البذرة العارية بشكل كامل تقريباً { حيث تغطى بصفيحة رقيقة جداً نصف شفّافة }. وهي البذرة التي نأكلها نحن وحيواناتنا، وكذلك هي المادة الأساسيّة لاستخراج زيت دوّار الشمس. وكما تعلم فكل مرّة تأكل بها بذرة دوّار شمس، فأنت تأكل جنين ثمرة نباتيّة مع كامل مخزوناتها الطاقيّة.  تنوعات لثمرة الفقيرة: 1دوّار شمس Helianthus annuus 2 ضرس الاسد Taraxacum sp ، 3 الدردار Ulmus campestris 4 ثمرة جنس Xanthium برّي مقابل المزروع. معاً ومطواعان يحتفظا دوّار الشمس المزروع والبرّي فيما بينهما بكثير من الفروقات: فلا يتمكّن النوع البرّي من تحقيق الإلقاح الذاتيّ، وساقه الرئيسيّ غنيّ بالفروع وكل فرع منها يقوم بانتاج الازهرار وهي بدورها مُختصرة ووافرة،وبذات الوقت ثمارها قزميّة بشكل مثير. من جانبه النوع المزروع، قادر على تحقيق الإلقاح الذاتيّ، يمتلك ساق رئيسيّ يفتقر للتفرعات وينتهي بمكان ازهرار رئيسيّ وتكون ثماره ذات قياسات كبرى. يمكن إجمال تلك الفروقات الواضحة في الجدول التالي:  جدول مقارنة بين دوّار الشمس البرّي والمزروع بالرغم من كل هذا الفارق بينهما، فإنّ كلاهما يشكلان ذات النوع Helianthus annuus وينموان سوياً في الولايات المتحدة والمكسيك، واحد في حقول الزراعة وآخر في حدودها. كذلك يُزهران بذات الفترة { بنهايات شهر مايو أيّار وصولاً لأكتوبر تشرين أوّل } ويتلقحان كلاهما بواسطة النحلوحشرات شبيهة. هذا يفترض حصول مشكلة، خصوصاً بالنسبة للزراعات المعدلة وراثياً لدوّار الشمس، باعتبار انهما يشكلان ذات النوع، فيتهاجن النوع البرّي والمزروع بسهولة هائلة. اضافة لأنّ تهجينات كتلك تكون خصبة بشكل كامل. وهنا تكمن إحدى مشاكل الدوّار المعدّل وراثياً، حيث لا يمتلك شيئاً يُفيد الصحة البشريّة، بل مع الزراعة البيئيّة: يمكن للدوّار " المعدّل وراثيا " العبور للجماعة البريّة من نوع مزروع، ما يؤدي لتغيير تلك الجماعة وبتبعات لا يمكن توقعها للنوع المتأثِّر ولاجل باقي النظام البيئي.  مقارنة بين دوّار الشمس المزروع، إلى اليسار،ودوّار الشمس البرّي إلى اليمين يُعتبر دوّار الشمس بالوقت الراهن أحد المصادر الهامّة لازهار الزينة والبذور في المعجنات المختلفة، لكن تبلغ أهميته ذروتها على المستوى العالمي: باستخدامه لانتاج الزيت النباتيّ الأكثر انتشار. تعود أهمية هذه البذور { سيما التنوعات الحديثة منها } على الصعيد الصناعيّلغناها الهائل بأنواع الدهون A.K.A. grasas وهي احماض دهنية غير مُشبعةونستهلكها نحن على شكل " زيت للمطبخ ". تتشكّل الثلاث احماض دهنية الرئيسية في دوّار الشمس، من: حمض زيت الكتان { 66.9% من كامل المحتوى الدهنيّ،حمض الزيتيك { 18.1% وحمض النخليك { 6.5% } ولو أنّ تلك النسب يمكن أن تتغيّر بحسب النوع، اضافة لانه لا يحتوي كوليسترول أبداً. كذلك يكون دوّار الشمس غنياً بالاملاح المعدنية مثل المغنيزيوم،الفوسفور والبوتاسيوم، كذلك يحتوي على فيتامينات مثل ب1 { التيامين }، ب3{النياسين }، ب6 وفيتامين إي، كذلك يُعتبر دوّار الشمس مصدراً لحمض الفوليك البالغ الاهميّة للحوامل. يبلغ انتاج زيت دوّار الشمس ما يكافيء 8% من الانتاج العالميّ من الزيوت النباتيّة، ويأتي قبله زيت الصويا { 55% }، زيت السلجم { 14% } وزيت القطن{ 10% } تشكّل روسيا البلد المنتج الاول له بحيث بلغ انتاج البذور بين العامين 2008 / 2009 حوالي 7 مليون طنّ، وبعد روسيا يأتي الاتحاد الاوروبي، أوكرانيا والأرجنتين.  أهم مُنتجي بذور دوّار الشمس في الفترة 2008/2009 وراثة معقّدة لماذا ندرس عملية التدجين؟ لا يشكّل التدجين بالنسبة لعلماء الأحياء مجرّد معرفة تطبيقيّة لتحصيل تنوعات جديدة، أنواع جديدة أو حتى أنواع نباتيّةجديدة مفيدة للاستهلاك البشريّ، بل يذهب الاهتمام بالدراسة لأبعد من هذا،حيث تسمح دراسة التدجين بامتلاك معلومة مُباشرة لتطور الانواع الحيّة. منذ أن أبدى داروين رأيه بهذا، بخلاف الكثير من معاصريه، من حيث أنّ كثير من النباتات المزروعة والحيوانات المدجّنة ما هي إلا نتاج انتقاء منذ أصلها البرّي الاقدم وليست نتيجة خلق إلهي، يرى اليوم علماء الاحياء في عملية التدجينحدث مثاليّ لملاحظة التشابك بين التغيرات الجينية { الوراثيّة}والتغيّر الموروفولوجي { التشكُّلي } للكائنات الحيّة، كذلك المجرى الزمنيّ الذي يمكن لتلك التغيرات ان تحصل خلاله. بُغية تحقيق عمل هامّ، يتحدث علماء الوراثة عن اعجوبة كبرى تسمى QTL .. لا حاجة للاصابة بالخوف!! فهذه الاحرف تشكّل اختصار لكلمات Quantitative Trait Locus حيث يشبه QTL مجموعة كتب تحتفظ كلها بالتصاق الكتاب بالآخر،وكلها بذات المكان، وكلها تتحدث عن ذات الموضوع وكلها تُشترى معاً { وليس فرادى أبداً}،بترجمة ما سبق إلى لغة بيولوجيّة: يشكّل QTL مجموعة من الجينات، كلها تُقيم جنبا لجنب، كلها تشغل منطقة محددة من الكروموزوم،كلها تساهم باعطاء ذات الملمح أو الميزة وكلها يتم توريثها كحزمة{ونادراً جداً على شكل منفصل }.  باستخدام QTL لدراسة الصلات بين الجينات والميزات،a نباتات مُشعِرة جداً وقليلة الشعر. b هجينة عن النباتات السابقة، c هجينة حتى الجيل الثامن منها،الخطوة التالية: تحديد المنطقة الجينية المُشتركة السهملكل النباتات { قليلة الشعر }. لقد عثرنا على المنطقة المتواجد فيها على الاغلب QTL المسؤول عن توليد " الكميات الكبيرة او القليلة من الشعر " تسمح دراسة مجموعات الجينات تلك بتحديد ارتباطها بميزة محددة { حجم الثمرة،اللون وشكل الازهار، فترة الإزهرار أو الإلقاح ... الخ }، أو القول بشكل معاكس، تسمح برؤية انّ عناصر جينيّة تقف وراء ملامح محدّدة أو ميزات في الفرد. تساعدنا هكذا دراسات على فهم كيف يمكن لبنية جينية للنباتات المزروعة باجتماعها مع البيئة، أن تساهم بتوليد تنوّع الخضار التي نراها اليوم. سندرس التدجين باعتباره تطوّر ينفذه الانسان، وجرى العمل بمستوى جينيّ لتوليد التنوع النباتي، والتي تُخبرنا حول أيّة مجموعات جينيّة قد كانت منتقاة خلال تلك العملية. يكونا نوعي النبات البرّي والمزروع مختلفين، حيث تكون الثمرة أكبر في النوع المزروع كقاعدة عامّة، التحكُّم بفترة النموّ والازهرار بما يناسب مصلحة المزارع،تفقد البذور قدرتها على الانتشار وتزداد قيمتها الغذائيّة،بذات الوقت تكسب النباتات القدرة على تحقيق الإلقاح الذاتيّ. يُعرف كل هذا باسم " متلازمة التدجين " ويكون مدروساً بسهولة من خلال QTL حيث من المعتاد أن تقف عناصر جينية وراء تغيرات هامّة في النباتات.لكن في حالة دوّار الشمس، هذه الآلية التي نجحت بدراسة الذرة، قد فشلت معه بصورة مثيرة. في العام 2002، نشر فريق بحث بقيادة عالم الوراثة والمختص بدراسة الفصيلة النجميّة Dr. John M. Burke مقالاً في المجلة العلمية Genetics والذي انتهى بإيراد خبر سيء للغاية. فقد امتلكت استراتيجيّة QTL لأجل كشف العلاقة بين علم الوراثة والنمط الظاهريّ fenotipo نتائج لا طائل منها وقدرة ضئيلة لتفسير الفروقات القائمة بين دوّار الشمس المزروع ودوّار الشمس البرّي.بشكل غير متوقّع، وُجِدَ QTL لنباتات بريّة قد ولّدت ميزات خاصة بالنباتات المزروعة، وكذلك QTL لنباتات مزروعة قد ولّدت ميزات خاصة بالنباتات البريّة. وبالنهاية أشارت QTL للكرموزوم LG06 بأنّه قد جرى انتقاء ميزات غير مرغوبة لاجل النوع المزروع، الأمر الذي كان بلا معنى.لماذا يتم اختيار شيء لا رغبة فيه؟ ما الذي حصل بالضبط؟ يبدو أنّ هناك أشياء عديدة بذات الوقت تحصل. وبخلاف أنواع حيّة أخرى، فإنّ QTL لوحده قد أمكنه تفسير قسم جيّد من الفروقات بين النوعين البرّي والمزروع، نجد أنّه في دوّار الشمس، لكل QTL قد امتلك قدرة ضئيلة جداً لتفسير فروقات كتلك. وبحسب الباحثين فإنّ هذه الظاهرة تعود لحصول تغيُّر تدريجي من النوع البرّي الى المزروع. وفق هذه الصيغة تعمل كثير من QTL كمجموعة لتعطي كنتيجة دوّار الشمس المزروع، حيث يكون إسهام كل واحد منها صغير جداً. لدينا بالجانب المقابل الكروموزوم LG06 المتميّز بامتلاك QTL متصلة بملامح غيرمرغوبة بدوّار الشمس المزروع،لكنه امتلك كذلك علاقة قويّة بإنتاج الزيت! بناء على نتائج تلك الدراسات، عاد الانتقاء لذاك الكروموزوم لفترة قريبةولهذا لا يكون مرتبطاً بعملية التدجين البدائيّة ونعم مع الاصدار الحديث من هذه النبتة كمصنع للزيت. سيكون هذا الضغط الانتقائيّ الحديث قويّاً بما فيه الكفاية لاجل دفع انتقاء QTL للآثار المرغوبة بشكل قليل .  وجها الكروموزوم LG06 تُشير الاشرطة لمكان QTL في الكروموزوم. اللون الاسود: أثر مرغوب، اللون الابيض: أثر غير مرغوب. إلى اليسار: احماض دهنية مختلفة.إلى اليمين:DFlr إزهرار Lvs ، عدد وريقات الساق الرئيسيّ SDia قطر الساق، H/B عدد الزهيرات / الساق، AcWd عرض ثمرة الفقيرة، AcWt وزن الثمرة، Ray عدد جذور الوريقات، Brn عدد الافرع الجانبيّة، RySz حجم جذور الوريقات، Hght الوزن خلال الازهرار لكن العلم يشكّل نشاط عقول قلقة من النادر أن تستسلم. ففي العام 2008نشر عالم الوراثة ودارس النباتات Dr. Mark A. Chapman والعامل حاليا في مكتب علوم النبات بجامعة اكسفورد / بريطانيا في واحدة من أهم المجلات العلمية المختصة بعالم النبات وهي The Plant Cell وثيقة قد أضاءتبالنهايةبشكل أكبر على هذا الموضوع. من التنبؤات الاساسيّة للدراسة البيولوجيّة للتدجين: هو امتلاك " الجماعات " المدجّنة لتنوّع جينيّ أقلّ مما تمتلكه الجماعات البريّة التي تشكّل أصلها. ويعود هذا لسبب اقتضاء التدجين لانتقاء الجماعات المقيّدة كل مرّة أكثر بأفراد { الذين لا يكونوا سوى حاملي الملامح المُوجبة ليكونوا مدجنين } يمثلون نسبة مئوية قليلة من التنوّع الاجماليّ للجينات الموجودة بكامل النوع الحيّ. أثبت بحث الدكتور Chapman وجود هذه الظاهرة في دوّار شمسنا، حيث أنّ جماعاته المزروعة قد كانت أكثر فقراً بالجينات من الجماعات البريّة. وذهب أبعد من هذا، حين أثبت بأنّ التنوعات " الأوليّة " لدوّار الشمس المزروع قد كانت أغنى جينياً من " الأحدث أو الحديثة ". بوقت لاحق تعرّض دوّار الشمس لضغط انتقائيّ سواء خلال فترة تحولها من النوع البرّي للنوع المزروع، كما في استمرار تطورها من اصداراتها المزروعة الأُوَلْ وصولاً للتنوعات الأكثر حداثة. وهذا ما تمّ تأكيده عبر دراسات أحدث تُشير لأنّ دوّار الشمس الحالي يحتفظ بما نسبته 67% من التنوّع الجينيّ لمقابله البرّي.  تنوّع جينيّ لدوّار الشمس البرّي نوع WILD والمدجَّنة البدائيّة primitivos والمدجَّنة المتقدمة avanzados ، تُشير الاشرطة للتنوّع القائم في كل مجموعة جينات قد تعرضت للانتقاء بفعل التدجين{الشريط الاسود }، والانتقاء بفعل انتقاء لاحق افضل { الشريط الرمادي}، ومن لم يتعرّض للانتقاء { الشريط الابيض } بكل نوع نباتيّ بخلاف الدكتور Burke فقد اعتبر فريق بحث الدكتور Chapman بأنّ التصوير الجينيّ:" لم يزوّد بأيّ معلومة مُباشرة عن الأنماط الظاهريّة المتأثِّرة بالجينات المعثور عليها تحت تأثير الانتقاء، وبهذا تمّ إحراز التقدّم نحو خطوة هامّة، هي اكتشاف أنّ جينات تعرضت للانتقاء بالرغم من أنّ وظيفتها لم تكن معروفة. تمّ التوصُّل لتلك النتائج عند تركيز البحث المُباشر بجينات مختلفة لدوّار الشمس البرّي، لدوّار الشمس " الأوّلي " وللتنوعات " الأحدث " منه. كان كل هذا بهدف مقارنة تنوّع الجينات في كل مجموعة ومحاولة استنتاج الضغط الانتقائيّ الذي أمكن لكل مجموعة التعرّض له، وفق هذه الصيغة تواجد ما يقرب من 36 جين مختلف. على الرغم من أنّ وظيفتها كانت مجهولة، فإنّ فريق الدكتور Chapman قد امتلك صيغتي نظر للدور الذي يمكن أن تلعبه تلك الجينات: الصيغة الأولى:البحث عن جينات معروفة متطابقة أو شديدة الشبه بتلك الجينات المجهولة. الصيغة الثانية: اكتشاف أين كان موضع الجينات المجهولة في الكروموزوم، الأمر الذي سيسمح بمعرفة أيّ QTL قد ساد عليها. لذلك وعندما تحرّوا عن طبيعة تلك الجينات، اكتشفوا أنها كانت تمتلك وظائف أكثر تنوّع، عوامل نسخ kinasas, transferasas ، بروتينات بنيوية. لا تظهر هذه بوصفها أسماء مُنتظَرة، فلا شيء عن " جين للبزر " أو " جين للفصوص الكبيرة ". لكن وعلى الرغم من الحاجة لوضعها قيد الدراسة العميقة للآن، فإنها وبشكل أوليّ جينات من المعتاد امتلاكها لوظيفة مُنظِّمة، ومن الممكن أن تتراوح من النموّ حتى التركيب لأيّ مركَّب. رغم هذا هناك قسم من " الجينات المنتقاة عبر التدجين " قد نتجت بوصفها ذات أهميّة خاصة، كانا جينان منهما متطابقان مع جينات معروفة مسؤولة عن تنظيم الإزهرار عند أنواع أخرى، اضافة لانها قد ظهرت في منطقة كروموزوميّة كان يُعرف بحضور تلك الجينات فيها مع ذات الوظيفة. من جانب آخر، امتلكت 4 جينات أخرى وظيفة لصيقة جداً بنموّ ثمار الفقيرة aquenios وبالبروتينات الحاضرة في البذور، نعم فنحن نتحدث عن البزر.  مقارنة بين اقراص دوّار الشمس البرّي والمزروع، في المركز نجد النوع البرّيوحوله نجد النوع المزروع لكن مع التجديدات الجينيّة { وراثيّة } والتغيرات التدجينيّة لكن لم تتمتع كل دراسات علم وراثة دوّار الشمس بكل هذا النجاح. لنذهب أبعد من عملية التدجين. ما هي الطريقة الأشهر لإدخال التجديدات الوراثيّة في الكائنات الحيّة؟ من المؤكّد انكم تعرفوها، وهي: إحداث الطفرات.وهذا ما يتعرّض لهجوم الحركة الخلقية الشديد. فهم يؤكدون التالي: تكون الطفرات سيئة، وتقوم بتسبيب التشوهات، المرض والموت فقط. وفي جزء من هذا الموقف، جزء فقط: لديهم الحقّ. فلا تمتلك غالبيّة الطفرات أيّ أثر، ومن القسم المتبقّي يكون معظمها ضارّ. وفقط نسبة مئوية منخفضة نسبياً تكون مفيدة. بالتالي الخلاصة المنطقية هي، من السهولة بمكان تحميل شيء بدل تغييره أو تحسينه بشكل جوهريّ. لكن يوجد طرق تجعل الاشياء أسهل، ويبدو أنّ الخلقيين إمّا أنهم لا يعرفونها أو يفضّلون عدم الاشارة لها. إحدى تلك الطرق هيالتضاعف الجينيّ. ويتأسّس التضاعف الجيني على تضاعف جين أو مجموعة من الجينات، أو يمكن أن تكون ثلاثيّة أو رباعيّة. وهذه الظاهرة تكون عامةوموثّقة في الطبيعة. فيما لو يتضاعف الجين، ما العمل مع النُسخ " الزائدة " ؟ فبينما يكون لدينا على الأقلّ نسخة من هذا الجين بكل تلك الاهميّة وظيفيا،يمكننا التحكّم بباقي الجينات كيفما نريد. في الواقع، تشكّل الجينات المكرّرة مادة ممتازة لاجل الابتكار التطوريّ، حيث يمكن للجينات الزائدة أن تصنع أشياء أخرى، من فقدان وظيفيّة ما { باعتبارها نسخة فلن يحدث شيء}،وصولاً لاكتساب وظائف اضافيّة، وظائف جديدة كليّاً أو جزئيّاً,أو تحقيق ذات الوظيفة ولكن بصورة فعّالة ودقيقة أكثر { بالتالي تنظيم هذا الجين سيكسب مستوى أكبر من العمق}.  تضاعف جينيّ. إثر حصول تضاعف كهذا، سيتكوّن الكروموزوم من نسختين لهذا الجين، ومن QTL أو من تلك المنطقة بالكروموزوم تمّ تسجيل حضور هذه الظاهرة في نبات دوّار الشمس. إنها مجموعة جينات CYCLOIDEA-like التي تمتلك دوراً في تشكيل الأزهار. تكون تضاعفاتها أقدم بكثير من تضاعفات عند الإنسان، حيث توجّب حصولها بدوّار الشمس منذ الفترة التي تعود إلى ما قبل 41 – 44 مليون عام.وقد تشكلت تسلسلاتها وفق الآتي: يمتلك CYC1 نسختين، يقوم HaCYC1 بتحقيق وظيفته في كل الانسجة ما عدا الجذور، و يعمل HaCYC1b في الجذور فقط، يمتلك CYC2 كثير من النُسخ،يعبّر HaCYC2b في كل الانسجة، فيما يعملان HaCYC2a و HaCYC2e في جميع مكونات الازهرار العنقودي، بينما يعمل HaCYC2d فقط في الازهار ويعمل HaCYC2c فقط في الازهار من نوع اللسينيات lígula ، كذلك تظهر 3 نسخ من CYC3 بالرغم من امتلاكها كلها دور متشابه جدا.بالنتيجة لقد تقدمت الجينات المضاعفة خطوة بالتخصّص، بحيث تعمل في أماكن محددة وليس بغيرها. أو تلعب جينات المجموعة FLOWERING LOCUS T-like (FT-like) دوراً هاماً في تنظيم عمليّة الإزهرار بحسب مدّة اليومبصورة تسمح للنبات بمعرفة فيما لو نكن بفصل الخريف – الشتاء { أيام بفترة نهار تقصر باستمرار } أو في فصل الربيع – الصيف { أيام بفترة نهار تطول باستمرار } حيث يهمّ النبات معرفة الحقبة للإزهرار باللحظة الأفضل. حسنا،تعرضت جينات دوّار الشمس للتضاعف في العصر الحديث، ولهذا تمتلك 4 نسخ لذات الجينHaFT1 العامل في البارض{النقطة التي سينمو منها الافرع والاوراق المستقبليّة }، يُعبّراHaFT2 و HaFT4 في الأوراق النامية لا في البارض، وتكون النسخة الاخيرة للجين HaFT3 غير عاملة.  مؤثرات الطفرة في الجين CYCLOIDEA ،الى اليمين: زهرة نوع برّي Linaria sp ، الى اليسار: ذات الزهرة لكن مع طفرة بالجين CYCLOIDEA و لمزيد من الظرافة: لقد كانت تلك الطفرة متماثلة جينياً epigenética وهذا بفضل الآلية الرئيسيّة له metilación للجين من الواضح أن مجموعة الجينات FLOWERING LOCUS T-like (FT-like) قد لعبت دورها هذا خلال عملية التدجين لدوّار الشمس. وعبر تقنيات مؤسّسة على قيممُبالَغ فيها للمُعامل QTL وجدوا بأنّ الجينات المتنكِّبة عملية تنظيم الإزهرار والمنتمية لعائلتيّ TF وFTL قد امتلكت كل الأوراق الدالّة على تعرضها للانتقاء خلال عملية تدجين هذا النبات. حيث تتنكّب تلك الجينات،بين وظائف أخرى، تنظيم الإزهرار وتعطي الانطباع بأنّ الضغط الانتقائيّ قد كان مُستثمراً خلال تاريخه. على سبيل المثال، يتركّز دور الجين HaFT1 بتأخير الإزهرار. ويظهر أنّه تمّ انتقاء هذا الجين خلال أوائل لحظات التدجين لدوّار الشمس، سواء بصورة مباشرة أو بصورة غير مباشرة كعاقبة لانتقاء ملامح أخرى مرغوبة يسببها هذا الجين. تؤكد دراسات أخرى محقّقة على جينات FT في نبات ينتمي للفصيلة الصليبيّة هو Arabidopsis أو في البندورة بأنّ هذه الجينات تقوم بإتمام دور متصل ببنية النبات ونموّ الأوراق، وفيما لو تقوم بذات الدور في دوّار الشمس، فهذا يشكّل سبب جيّد لانتقائها. من جانب آخر، يبدو أن الجين HaFT1 قد تعرّض لضغط انتقائيّ سلبيّ بأزمنة حديثة، ما يعني انتقاء هدفه الغاء الجين HaFT1 أو الغاء آثارهبحسب الرغبة بالحصول على نبات إزهرار باكر من قبل بعض المزارعين.  حقل دوّار شمس يأتينا جانب هام آخر من أعمال الدكتور Burke والذي كان حتى الآن بحالة انتظار: كنتيجة لدراسة محققة في العام 2002 ومؤسّسة على QTL ، لكن لم تكن كل نتائج تلك الدراسة من النوع المثير، فلقد عثروا على 14 QTL مرتبط بصورة أو بأخرى بنموّ البذور. لقد ارتبطوا بنموّ الطول، العرض والوزن للثمار{البذور }، وكما هو مُنتظَر في التحوّل من النبات البرّي إلى النوع المزروع.نشر ذات الباحث بعد مرور 3 سنوات معلومات عن تنوعات QTL مرتبطة بمحتوى الثمرة من الزيت. حديثاً وعندما تمكّن الكائن البشريّ من تحويل دوّار الشمس لآلة مُنتجة للزيت، ويبدو أنها ظاهرة تعاكس ما رأيناه حتى الآن، فقد شكّلت حادثة أثّرت بقليل من QTL ولكن بتبعات كبرى لكل واحد منها.  مقارنة بين ثمرات دوّار الشمس المزروعة { الى اليسار } والبريّة { الى اليمين } ملاحظة QTL الواردة هنا قد وردت أعلاه، وهي اختصار لتعبير مكوّن من 3 كلمات فحواها حيث يشبه QTL مجموعة كتب: تحتفظ كلها بالتصاق الكتاب بالآخر،وكلها بذات المكان، وكلها تتحدث عن ذات الموضوع وكلها تُشترى معاً { وليس فرادى أبداً }. بترجمة ما سبق إلى لغة بيولوجيّة: يشكّل QTL مجموعة من الجينات، كلها تُقيم جنبا لجنب، كلها تشغل منطقة محددة من الكروموزوم، كلها تساهم باعطاء ذات الملمح أو الميزة وكلها يتم توريثها كحزمة { ونادراً جداً على شكل منفصل}. كلام مذكور سابقاً في هذه السلسلة. يكون تحوّل دوّار الشمس لمصدر للزيت موضوع هامّ. ففي هذا العام 2012، تمّ نشر بحث علميّ مُحقّق بشكل مُشترك بين اختصاصيّي علم الوراثة، التطور والتدجين لدوّار الشمس للدكتورين Burke و Chapman. يتحقق المسار البيوكيميائيّ المسؤول عن تركيب الاحماض الدهنيّة المتراكمة في بذور دوّار الشمس من خلال سلسلة من الجينات FA ، حيث تُثبت هذه الدراسة ومن جديد فقدان التنوّع الجينيّ بهذه المجموعة من الجينات، وهذا متوقّع كعاقبة للتدجين. أحد تلك الجينات لم يكن منتقى من قبل الانسان، لكنه منتقى بصيغة طبيعية وهو FAD7 والذي يظهر أنّ انتقاؤه قد حدث في اللحظة التي جرى بها انقسام Helianthus annuus و Helianthus petiolaris لنوعين مختلفين. من جانبهما، أبرزا الجينان FAD2-3 و FAD3 عملية انتقاء خلال اوائل لحظات التدجين لدوّار شمسنا الراهن، بيّنت باقي الجينات المدروسة FAB1, FAD2-1, FAD6 و FATB حصول انتقاء فقط خلال حقبة تالية لتدجينهاعندما شاعت زراعة دوّار الشمس. أوائل الجينات FA المنتقاة بفضل التدجين، هي FAD2-3 و FAD3 المُرتبطان بتحويل حمض الزيتيك ácido oleico إلى حمض زيت الكتّان ácido linoleico هكذا كتحوّل هذا الأخير إلى linolénicoبشكل متتابع. من المعروف أن تلك الاحماض الدهنيّة تؤثّر سواء على استساغة الاطعمة كما على الإنتاش وكلاهما سببان منطقيان لانتقاء هذه الجينات. هل فرضت الاستساغة معدّل إنتاش محدّد؟ أو سبّب الإنتاش تغيُّر بعملية الاستساغة؟ أو كلاهما معاً؟ من الصعب الإجابة. من جانبها تمّ انتقاء باقي الجينات مباشرة لاجل انتاج الزيت من البذور.  انتاج زيت دوّار الشمس وفق الآلية هذه أصول النوع المزروع في الفترة التي تعود لما قبل 11000 – 5000 عام، بدأت جماعات بشرية بمختلف أنحاء العالم بتدجين أنواع نباتيّة وحيوانية مختلفة، وبهذا تمّ تسجيل بداية انتاج الغذاء وتكوين اقتصاد جديد وتغيير جذريّ بحياة الكائن البشريّ وبشكل دائم. ولحسن الحظّ أن نوعنا لم يكن قد ابتكر البنوك بوقتها، ما أفسح المجال للبقاء على قيد الحياة بهذا العالم القاسي. يسمى الانتقال من حياة الرعي والصيد والتقاط الثمار إلى حياة استقرار وزراعة بالثورة النيوليتيّة Neolithic Revolution ، وحولها نتكلم وفق تفاصيل مؤكّدة في موضوعنا هذا من سلسلة البستان التطوريّ، حيث انّ تلك الحقبة قد كانت محطّ بحث عميق في علم الأحياء وعلم الآثار خلال القرن المُنصرم. تُعتبر شمال أميركا من أهم النقاط التي أصّلت للزراعة بشكل مستقلّ في الكوكب الارضيّ. فلقد زُرِعتْ أنواع نباتيّة لم تكن معروفة بأوروبا، مثل marshelder { Iva annua } و huauzontle { Chenopodium berlandieri } وعائلات أخرى مثل اليقطين Curcubita pepo ودوّار الشمس Helianthus annus وهو بطل موضوعنا هذا.  نوعين مدجنين في اميركا الشماليّة القديمة, الى اليسار: نوع marshelder - Iva annua ، إلى اليمين: نوع huauzontle - Chenopodium berlandieri يتوزّع دوّار الشمس البرّي على نقاط شاسعة من القارة الأميركيّة، تمتد من وسط وغرب الولايات المتحدة الاميركية وصولاً لجنوب شرق كندا وشمال المكسيك.ممكن أن تكون أسباب الاهتمام به بوصفه مصدر غذائيّ، ومواد ملونّة وطبيّة.عُثِرَ على الدليل الأكبر لحضور دوّار الشمس في الولايات المتحدة عبر مجموعة متفحمة مكونة من 6 بذور في منطقة Hayes (Tennesse) وبفضل الكاربون 14 تمّ تأريخها بحدود ما قبل 4260 ± 60 عام، يسجّل علماء الآثار أنها مزروعة { وليست بريّة } لانّ طول الواحدة منها يتجاوز 7 ميليمتر وهو الحجم الأقلّ المُقترح بحالة دوّار الشمس المزروع. بالرغم من وضع تلك الطبيعة التدجينيّة بحالة شكوك، فإنّ دراسات جرت بوقت لاحق وأثبتت بأنّ حجم وطبيعة تلك البذور المعثور عليها بمنطقة Hayes تتجاوز النوع البرّي بوضوح شديد. بذات الحقبة ما قبل 4500 و 4600 عام، كذلك عُثِرَ على بقايا دوّار شمس مدجّنة بمنطقة San Andrés في ساحل خليج Tabasco في المكسيك، وتتراوح أبعاد ثمارها من 7.8 . 4.4 ميليمتر إلى 8.2 . 4.5 ميليمتروهو حجم أكبر بكثير من حجم ثمار النوع البرّي والذي يُعثر عليه بذات المنطقة. مع هذا تعرّضت تلك العيّنات للنقد، والذي تركّز على التشكيك بطبيعة صور البذور ذات النوعيّة السيِّئةوضرورة الحاجة لصور ميكروسكوبيّة الكترونية للبتّ بالأمر وتحديدها بوصفها ثمار دوّار شمس. بالنتيجة بقي حضور دوّار الشمس المرزوع في المكسيك بتلك الحقبة بحالة " خطر ". تكون تلك الاعتراضات مألوفة علمياً. فالبراهين، الأدلة المضادّة أو كشف ضعف الأدلّة المُقدّمة، تكون أمر معتاد. والردود، ما لم تكن هدّامة، من المألوف أن تتناقض مع ردود مدعومة بالأدلة الأقوى والمعطيات. هكذا ورد ردّ على التأكيدات المُشار لها أعلاه أثبت حضور دوّار الشمس المرزوع في المكسيك منذ ما يزيد على 4500 عام، حيث اكتُشِفَتْ ثمار جديدة في كهفCueva del Gallo بالمكسيك بحال حفظ ممتاز اضافة لتضمّن المقال المنشور حول هذه اللقى " لصور ميكروسكوبية الكترونية اثبتت بما لا يدع مجال للشكّ انها ثمار دوّار الشمس".  الى اليسار، ثمرة دوّار شمس معثور عليها بمنطقة Cloudsplitter Rockshelter ولاية كنتاكي، الى اليمين، ثمرة دوّار شمس معثور عليها في Cueva del Gallo بالمكسيك تمّ تحقيق هذا العمل من قبل فريق بحث علمي برئاسة عالم أحياء وآثار ونباتات David L. Lentz بمعهدMcMicken College ، دعم الفريق بحثه من خلال دراسة لغويّة قد أثبتت بأنّ هنود المكسيك، شعوب los Mayo, los Tzotzil Maya و los Zapotec قد استخدموا أسماء محدّدة وحيدة لتحديد دوّار الشمس،وجماعات هندية أخرى { كشعوب Huastec, Mixe, Nahua, Otomi, Popoluca, Raramuri / Tarahumara, Seri, Tepehuan, Totonac, Tzeltal و Zoque قد استخدموا اسم مختلف صوتياً بشكل كبير عمّا يستخدم الاسبان. بكل الاحوال هذا يقدّم لنا الدليل على معرفة دوّار الشمس عند تلك الشعوب قبل مجيء الغزاة الاسبان. هكذا نجد أنّ دوّار الشمس يمتلك مؤشرات جزيئية، أثرية ولغوية تتسجّل على منطقة شرق اميركا الشمالية كمركز تدجين من جهة، ومن الجهة الأخرى توجد ذات المادة صالحة في المكسيك مُظهرة امكانيّة التدجين لدوّار الشمس في هذا الجزء من العالم. يتساءل العلماء: هل تمّ تدجين دوّار الشمس مرتين في مكانين مختلفين؟ أو في الواقع قد تمّ تدجينه مرّة واحدة؟ وفي حال كان الأمر هكذا، أين تمّ الأمر وكيف يصل من مكان لآخر ؟؟ مزيد من الأسئلة الصعبة. دراسة مؤسّسة على نماذج بريّة ومزروعة سواء في أميركا الشمالية أو في المكسيك، حاولت الاجابة على تلك الاسئلة من خلال علم الوراثة. وبحسب نتائجها، فإنّ الانواع المزروعة راهناً من دوّار الشمس في المكسيك تكون متحدّرة من جماعات بريّة من دوّار شمس أميركا الشماليّة. وربما حصل هذا الإدخال لهذه الجماعات منذ آلاف الأعوام، مع هذا لا يكون ممكناً استبعاد حصول تدجين مستقلّ في المكسيك لدوّار الشمس ويكون ممثليه منقرضين بوقتنا الراهن. وربما يعود سبب هذا بجزء منه للغزو الاسبانيّ للمكسيك وحصول تدمير لدوّار الشمس كرمز لشعوب مقيمة مثل Aztecas وثقافات أخرى بأميركا الوسطى.  حجم مُقارَن لبذور دوّار شمس معثور عليها بحقول تنقيب مختلفة، A بمنطقة Hayes بالولايات المتحدة الاميركية، B بذرة بريّة، C بذرة معثور عليها بمنطقة San Andrés بالمكسيك، D بذرة حديثة مزروعة. أبناء دوّار شمسنا: تطور على جناح السرعة لا يمكن ترك موضوع دوّار الشمس، دون الاشارة لقضيّة ظهور " نوع جديد " وتكيُّف مع بيئة مختلفة تماماً وقاتلة. وهذا يشكّل قضيّة أخرى من نشوء الأنواع، من تكوين نوع حيّ جديد، والذي يشترك بداية بعملية التهجين. لكن بخلاف ما رأيناه بحالة القمح Raphanobrassica sp والقطن، فلا نتحدث هذه المرّة عن هجائن عقيمة و " الطفرة الكبرى " المسماة poliploidía والتي تحوله لنوع نباتيّ جديد خصيب وقابل للحياة، بل نتحدث عن نباتات مُعتبرة أنواع حيّة مختلفة من قبل علماء النبات، والتي بتهجينها تقوم بتوليد نبات قابل للحياة لكنه مختلف جداً عن الأنواع التي شكّلت أصله. يا له من شأن صعب اعتبارها جميعها تنتمي لذات النوع الحيّ !!!!! بوقت ما انتشرت انواع دوّار الشمس السنويّة والبريّة بالقارة الأميركية.النوع El Helianthus annuus البرّي الذي أعطى الاصل للنوع المزروع، وهو نوع يعيش بتربة ذات تركيب طينيّ جزئياً، وهي تربة جيدة لاحتجاز الرطوبة وتركيز ملحيّ {24±10 جزء بالمليون }. ونحن البشر نعتبر هذا النوع من التربة مثاليّ لأجل زراعاتنا. من جانب آخر، يكون النوع Helianthus petiolaris كنوع برّي آخر هامّ، ويستوطن مناطق أكثر جفاف وذات طبيعة رمليّة وذات ملوحة { 120±27 { جزء بالمليون.  الى اليسار، نوع Helianthus annuus برّي، الى اليمين، نوع Helianthus petiolaris بوقت لاحق عُرِفَ نوع Helianthus paradoxus كنوع برّي يعيش فقط في مناطق ذات تربة مالحة، بحرية وتربات أخرى مالحة حيث يكون مؤشّر الملوحة 2.000 إلى 20.000جزء بالمليون!! حيث لا يمكن لنوعيه البرّي السابقان العيش في هكذا تربة ببساطة حيث يواجهان الموت المحتّم. حيث يمتلك كل نوع منها معدلات قبول للملح مختلفة، لهذا يمكنهم العيش ببيئات مختلفة دون الحاجة للتنافس فيما بينهم. حتى الآن أفترض أنّ كل ما مرّ منطقيّ، لكن هل تصدقون لو أخبركم بأنّ نوع H. paradoxus هو في الواقع الإبن الغير شرعيّ للتهجين بين النوعين H. annus H. petiolaris و ؟؟ لا يُشكّل نوع H. paradoxus حالة تساهل مع البيئات المالحة أكثر من قدرة أبويه،بل كذلك يقوم بتطوير أوراق عُصاريّة { غنيّة بالماء } بنسبة كبيرة ويمتلك قدرة كبرى على تخزين منسوب مركّز هائل من الملح في أوراقه، ويملك كلا التكيفين للبقاء على قيد الحياة في بيئات مالحة مقارنة بأبواه.والآن لدينا التساؤل، لماذا يحصل هذا طالما أقربائه من الأنواع الأخرى أقلّ تساهلاً مع الملح ؟ هنا تحضر النتائج غير المتوقّعة من تآلف جينات محدّدة والقدرة على الانتقاء. لاجل ذلك حقّق علماء أبحاث ذات طبيعة مختفة تجارب مخبريّة على بيئات مختلفة الملوحة، اختبارات في البيئة الطبيعية ودراسات جينيّة.لقد استخدموا أشطاء { ساق نباتيّة ثانوية } لأنواع H. annuus, H. petiolaris, H. paradoxus وهجائن* بين H. annuus و H. petiolaris ولاحقاً استزرعوا أشطاء لكل نوع سواء في دفيئة مختبرية كما في سكن طبيعيّ لنوع H. paradoxus بهدف دراسة كيفة ترتيبها خلال 46 يوم لاحقة. * ملاحظة: تمّ توليد تلك الهجائن من خلال أول تهجين بين H. annus و H. petiolaris. ويشكلان نوعان مختلفان، ولهذا تمّ تحصيل عدد قليل من الهجائن القابلة للحياة، ولهذا السبب بالذات تمّ تهجين متحدرها من جديد مع H. petiolaris لاجل إحراز عدد أكبر من الثمار الصالحة.  عيّنة ظريفة من نوع Helianthus paradoxus كانت النتائج هامّة. الاختبار ضمن حقل مفتوح، تمكّن فقط النوع H. paradoxus من عبور الاختبار { كان النوع الوحيد الذي بقي على قيد الحياة}وتمتعت الانواع الهجينة بقدرة متوسطة مقارنة بالأنواع الأبوية لتحمّل الملوحة وماتت كذلك. مع هذا أوضحت الدراسات الجينية بأنّه في النوع H. paradoxus يتم انتقاء QTL المرتبطة بالتحكُّم بالأملاح في النبات والبقاء على قيد الحياة في بيئات مالحة. كذلك كانت مجموعات الجينات تلك حاضرة في الانواع الهجينة، ولكن لم تتآلف بذات القدر في كل الحالات، فقد اظهرت اغلبية الهجائن حالة متوسطة بين النوع H. annuus والنوع H. petiolaris. لكن بعضٌ من تلك الهجائن قد بيَّنت ميزات غير حاضرة بأيّ من الأنواع الأبويّة وكانت حصريّة بالنوع H. paradoxus. من بين تلك الميزات المتفوقة عمّا لدى الأبوانالنموّ باحتفاظ الاوراق بالمياه، تحسُّن القدرة على تمثيل الكالسيوم من الوسط المحيط وتآلف هذه القدرة مع امكانية رفض أملاح أخرى أخطر على حياة النبات. ستشكل تلك الهجائن الجاهزة للعيش ببيئة مالحة متوسطة الأسلاف الممتازين للنوع H. paradoxus واعتباراً من تلك اللحظة ستدخل بإطار الانتقاء الطبيعيّ لإتمام القدرات الجديدة. لا يكون مفاجئاً هذا التمايز بالهجائن، فبمجرّد تحقيق التهجين، يُعاد تنظيم الجينوم، " تنوعات " مختلفة للجينات ذاتها،تسمى أليلات، تتآلف بطرق مختلفة وتعطي المجال لامكانات عديدة. فلا يكون غريباً أن يشكّلا النوعان Helianthus annuus و Helianthus petiolaris الأبوان للنوع Helianthus paradoxus فقد ساهما بظهور هجائن غير شرعيّة أخرى، مثل النوع المُثير Helianthus anomalus الذي يقطن في كثبان رمليّة،كذلك النوع المقدام Helianthus desertícola القاطن في الصحاري. كلاهما نوعان قد استوطنا بيئات قاسية مقارنة ببيئة الأبوين.  الى اليسار، نوع Helianthus desertícola ، الى اليمين، عيِّنة من نوع Helianthus anomalus نختتم موضوعنا هنا. يُعتبر دوّار الشمس من أكثر الانواع المزروعة عالميا، حيث يشكّل النوع الرابع من حيث الأهميّة لانتاج الزيوت النباتيّة ومصدر هامّ للغذاء لحيواناتنا المنزلية. تكون الفروقات بين النوع المزروع والبرّي منه واضحة، مع ذلك، ابرزت دراسات أن عملية تدجينه قد كانت مختلفة عمّا كان مُنتظراً، فلم يحصل انتقاء بضع جينات قليلة لاحداث تغييرات كبرى،بل تمّ انتقاء مجموعات كثيرة من الجينات لتحقيق تغييرات طفيفة معتدلةوهذا ساهم " بتأخير " واضح لدراسة دوّار الشمس مقارنة بأنواع مزروعة أخرى،ورغم هذا فقد تمّ الكشف عن انتقاء جينيّ بفضل تعديل الإزهرار، حجم الثمار وانتاج الزيت. ويُعتبر مفارقة أننا ختمنا المقال مع حالات تطوّر سريع في بيئات قاسية ومميتة أحياناً. أخيراً، دوماً هناك نقص بالبحث والدراسات،يُنتظر إكمالها دون أن تنتهي !!!! يتبع |
|
|
|
|
|
|
08-11-2016, 06:07 PM
|
رقم الموضوع : [9] |
|
عضو ذهبي
|
البُستان التطوريّ {9}: تطور البطّيخ  ربما سمعت غالبيتنا، كما تمتعت هذه الغالبية بفاكهة البطّيخ الأحمر { ويوجد ألوان أخرى }. يمتلك البطّيخ الأحمر تسميات عديدة { في الإسبانية Sandía }، حيث نجد بويكيبيديا أسماء له مثل: patilla، paitilla أو ببساطة " بطيخ الماء melón de agua " وهو اسم معروف بلغات أخرى. ويسميه الناطقون بالانكليزية watermelon. أما في كاتالونيا فيسموه síndria، síndia أو meló d’Alger، حيث تكون Alger اشارة لعاصمة الجزائر. ويسموه ببعض المناطق الحدودية بين الإكوادور والبيرو sandilla. بحسب القاموس الآكاديمي الملكيّ الإسباني، تعود كلمة Sandía للأصل العربي الاسباني sandíyya والذي بدوره يعود للمُصطلح العربي الفصيح sindiyyah المبنيّ على تعبير " من السند " أي ينتمي لمنطقة السند الواقعة في الباكستان اليوم، وكما ترون فحتى اللغة تتطوّر!  بطيخ للجميع. ثمار كاملة وأخرى مُقطّعة. لاحظوا اللون الأحمر المميز لبطيخنا المزروع بناء على بيولوجيّة البطيخ الأحمر، فهو يكون عضو في رتبة القرعيّاتCucurbitaceae، وتضمّ هذه الرتبة 118 جنس و825 نوع موصوف. وتضم هذه الرتبة كثير من الانواع الأخرى الشهيرة مثل البطيخ الأصفر Cucumis melo L، الخيار Cucumis sativus L، اليقطين Cucurbita maxima Duchesne، الكوسا Cucurbita pepo L بين أنواع غيرها. تتمدَّد معظم تلك الأنواع خلال نموّها على الأرض أو على دعائم تُوضَع لهذا الغرض. يمكن لهذه النباتات أن تعيش لسنوات عديدة perennes أو تعيش سنوياً { من حالة البذور الإنتاش، النموّ ووصولاً لانتاج البذور الجديدة حيث يمرّ عام كامل أو أقلّ بقليل ثمّ تموت النبتة }. ومن العتاد أن تكون أزهارها ذات جنس وحيد أزهار ذكريّة تقوم بانتاج الأسدية { جمع سداة } فقط وأزهار إناث تقوم بإنتاج البُنى التي تقود إلى البويضات { estigma ، estilo المدقّة والمبيض }. رغم هذا فيمكن لذات النبتة إنتاج أزهار ذكريّة وأزهار أنثوية. كما أنها تحتاج لنشاط الحشرات { سيما أنواع عديدة من النحل } لكي تتمكّن من تحقيق الإلقاح.  إلى اليسار زهرة ذكر بطيخ. إلى اليمين زهرة أنثى بطيخ يمثّل انتاج البطّيخ الأحمر ما نسبته 7% من الإنتاج النباتيّ العالميّ، حيث يتم انتاج حوالي 90 مليون طنّ سنوياً منه. وتكون بلدان إنتاجه الرئيسيّة الصين، تركيا، إيران، الولايات المتحدة الأميركية ومصر. والصين لوحدها تُنتج نصف الانتاج العالميّ تقريباً، كما انّ الصين وتركيا اكثر بلدين يخصصا اكبر مساحة لهذه الزراعة عالمياً. يُزرع البطيخ الأحمر بصورة رئيسيّة لأجل ثماره والتي يوجد منها تنوُّع بالأشكال، الأحجام، الألوان والطعومات. ومن المثير أنّ Lloyd Bright من ولاية آركانساس الأميركية قد سجّل رقم قياسيّ غينيس بفضل وزن بطيخته الذي بلغ حوالي 121.93 كيلو غرام في العام 2005. كما أنّ البطيخ الأحمر من الفاكهة الصحيّة، حيث يبلغ محتواه من المياه نسبة 90-92% وهو غنيّ بالفيتامينات آ وسي، وكذلك غنيّ بالليكوبين الذي يمنحه اللون الأحمر اضافة لكونها مواد مضادة للسرطانات، اضافة لجزيئات أخرى لها تأثير صحّي على القلب البشريّ.  مُزارع مع بطيخته العملاقة. لا أعتقد أنها فوتوشوب! يوجد في الولايات المتحدة الأميركية جمعية خاصة بزراعة البطيخ وتقديم الجوائز لمحققي أرقام قياسية بأحجامها، لدى الأميركيين هاجس غريب يتعلق بالأحجام! حين نتحدث علمياً عن البطيخ الأحمر الذي نزرعه، فسيكون اسمه الحقيقيّ Citrullus lanatus بحسب Matsum. & Nakai 1916. ويعود أصل Citrullus للمُصطلح اللاتيني / اليوناني citrus وهذا ما لا أفهمه حيث أنّ citrus هو الاسم العلميّ للبرتقال والليمون. اما lanatus فهو مُصطلح لاتيني معناه صوفيّ lanudo وهذا بفضل كميّة " الوبر " القائم في سيقان وأوراق النبات. لكن البطّيخ Citrullus lanatus لا يكون العضو الوحيد في الجماعة. ففي الواقع يشكِّل الجنس Citrullus مجموعة صغيرة ينتمي لها 4 انواع من النباتات وكلها تعيش بمناخ صحراوي وأصلها أفريقيّ. تكون تلك الأنواع هي على التوالي: - Citrullus lanatus وهو الذي ينتمي له بطيخنا الأحمر المزروع كما أشرنا سابقاً. وهو نبات سنويّ زاحف تمتدّ سوقه على الأرض. - Citrullus colocynthis L. Schrad وبخلاف بطيخنا الأحمر فهذه النبتة تعيش لعدّة سنوات perenne. لكنها تُزرَع كذلك اعتباراً من شمال افريقيا وصولاً للجنوب الشرقي الآسيوي، وهناك من يعزو لها فوائد طبيّة. - Citrullus ecirrhosus Cogn عبارة عن بطّيخ برّي متوطِّن في صحراء ناميبيا ويعيش لعدّة سنوات perenne كذلك. - Citrullus rehmii De Winter بطّيخ برّي متوطِّن في صحراء ناميبيا ويعيش لسنة واحدة فقط { دورة حياته سنوية }. ويمكن رؤية أشكال مختلفة لبطيخنا الأحمر Citrullus lanatus. ويرى بعض العلماء فيها تنوعين هامين فقط: 1- Citrullus lanatus var. Lanatus وهو بطيخنا الأحمر المزروع بمستوى عالميّ ويمكن رؤيته في الأسواق. 2- Citrullus lanatus var. Citroides ويتركّز زرعه في جنوب أفريقيا وآسيا { بالرغم من حضوره حاليا بقارات أخرى } والذي يُعتبر كسلف مُحتمل لبطيخنا المزروع والذي يضمّ عدّة جماعات بريّة. يكون الأمر مختلف لدى علماء آخرين، حيث يرون وجود 3 أنواع فرعيّة { وليس تنوُّعات } مختلفة هي: 1- lanatus الذي يُعتبر من أقدم الزراعات لهذه النبتة والمُكافيء للنوع var. Citroides. 2- mucosospermus والذي يمثّل نوع بطيخ اسمه Egusi والمزروع بأفريقيا فقط. 3- vulgaris والذي يمثل " بطيخ الصحراء الأحمر " وينتمي لنوع بطيخنا الأحمر المزروع. مع هذا، ما هو تاريخ البطيخ الأحمر؟ كيف أثَّر التطوُر به؟ يكون هذا قصّة أخرى.  حقل بطيخ مُعتنى به، نوع بطيخه Citrullus lanatus var. Lanatus الماضي الجينيّ التائه يُعتبر ماضي البطّيخ الأحمر مُشوّشاً. لكن سنحاول جلاءه. ولحسن الحظّ ففي شهر أوغسطس / آب 2013 المُنصرم جرى التوصُّل لنتائج بحثية حول هذا النبات، فقد نشرت مجلة الطبيعة المرموقة Nature أوّل مُسوَّده للجينوم الكامل للبطيخ الأحمر وهو عمل شاق حقّقه عدّة مراكز بحث في الصين وفي الولايات المتحدة الأميركية والدانمارك. Guo, S. et al. 2013. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nature Genetics 45: 51-58. يكون جينوم البطّيخ الأحمر صغيراً بالمقارنة مع جينومنا البشريّ، فهو يضمّ 11 زوج كروزومات فقط، بينما نحن نمتلك 22 زوج كروزومات. ويكون حجمه بحدود 425 مليون زوج قاعديّ، بينما يكون حجم جينومنا نحن بحدود 3000 زوج قاعديّ. لكن هذا لا يشكّل تقليل من الأهميّة، فيبلغ حجم جينوم القمح 16000 مليون زوج قاعديّ. بمعنى تتفوّق علينا كبشر عشبة كالقمح .. تخيّلوا!! من أهم نتائج هذا العمل البحثيّ، تأكيد أنّه بزمن محدّد قصيّ جداً وعندما لم يكن البطّيخ الأحمر هو ما نعرفه اليوم، حدثت ظاهرة قد ورثها كل المتحدِّرين، هي عمليّة تضاعف ثلاثيّ للجينوم { يشكّل الجينوم كامل المادة الوراثيّة / الجينيّة في النوع الحيّ }. هذه ليست قصّة جديدة، فلقد تحدثنا بهذه السلسلة { الجزء الثاني: تطوُّر العنب } منذ بضع سنوات عن شيء شبيه. لكن حسناً، قد مضى زمن طويل على هذا، وتقوم دراسات جديدة بتأكيده ولهذا لما لا نعيد التذكير به؟ عندما جرى في العام 2007 نشر جينوم شجرة العنب، جرى اكتشاف حدوث تضاعف ثلاثيّ بمناطق كبيرة بجينومها. وكان كِبَرْ حجم تلك المناطق الجينوميّة كافياً لإطلاق الفرضيّة التي تقول: هل كان ممكناً حصول تضاعف جينومي ثلاثي عند السلف الأبعد للدوالي { أشجار تُعطي ثمار العنب }؟ وبوقت لاحق بحث الباحثون المناطق المتضاعفة ذاتها عند ثلاث أنواع نباتيّة أخرى هي على التوالي: حور كاليفورنيا Populus trichocarpa، نبتة arabidopsis التي تكون مفضّلة عند أخصائيّ البحث الجينيّ الوراثيّ Arabidopsis thaliana والأرُّز Oryza sativa L. وقد كانت نتائجهم مثيرة للاهتمام جداً، فقد امتلكا حور كاليفورنيا ونبتة arabidopsis ذات التكرارات، لكن لم يمتلك الأرُّز هذا الأمر.  وضع المناطق المتضاعفة ثلاثيّاً { الاشارة لهذا بألوان مختلفة } في كل واحد من الصبغيات / الكروموزومات لشجرة الكرمة a ، لحور كاليفورنيا b ولنبتة arabidopsisc، وقد لا يكون واضحاً كفاية التضاعف الثلاثيّ عند الحور ونبتة arabidopsis، لكن يجب الانتباه لوجود اختلاطات في التسلسلات بتلك الأنواع. ويُعتبر جينوم الكرمة هو الأقرب للسلف بالتالي اقترحوا فرضيّة تقول: إثر انفصال سلالات أحاديّات الفلقة وثنائيّات الفلقة { منذ 130 إلى 240 مليون عام } سيحدث تضاعف جينيّ ثلاثيّ عند مجموعة صغيرة من النباتات المنتمية لسلالة ثنائيّات الفلقة، ستكون هذه النباتات ذات المادة الجينية الثلاثيّة التضاعف السَلَفْ المُشْتَرَكْ لكل النباتات التي نعرفها بيومنا هذا مثل الوردانيّات { التي تشتمل على حور كاليفورنيا، arabidopsis والكرمة بين كثير من الانواع النباتيّة الحيّة الأخرى }. فيما لو كان هذا العمل صحيحاً، وفيما لو يكن فهمنا للتطوُّر صحيح، كان يجب إكمال نبوءة كانت قائمة: بحيث أنّ البطّيخ الأحمر يكون بحسب الدراسات النباتيّة عضو في مجموعة الوردانيّات، ويجب أن يمتلك ذات السلف المُشترك ذو الجينوم ثلاثيّ التضاعف الذي تمتلكه الكرمة، arabidopsis وحور كاليفورنيا، بالتالي فيما لو ندرس جينوم البطّيخ الأحمر سنتمكن من رؤية أجزاء كبيرة من مادته الجينية / الوراثيّة ذات تضاعف ثلاثيّ. ملخّص نشوئيّ تطوريّ لظهور سداسيّ الصبغي الأصليّ. تمثِّل النجوم حوادث التضاعف الجينيّ حسناً، جرى تحقيق سَلْسَلَةُ جينوم البطّيخ الأحمر هذا العام 2013 وهو يؤكِّد تلك الفرضيّة { وعملياً يؤكّد فهمنا المُقترب من " تطوُّر الأنواع الحيّة " }. وفق هذا العمل فقد حدث التضاعف الجينوميّ الثلاثي الكامل لسلف الوردانيّات في حقبة متموضعة بين 76 – 130 مليون عام خلال عصر الكريتاسيّ { عندما سادت الديناصورات وزواحف أخرى في كوكبنا }. ولا يشكِّل هذا الأمر ظاهرة نادرة، ففي الطبيعة تشكِّل التضاعفات الجينية نوع من الطفرات شديد الخصوصيّة ويحدث هذا في عالم النباتات بصورة أكبر مما كان يُرى سابقاً. بصيغة مضت عبرها النباتات القديمة من امتلاك زوج مجموعات صبغية لامتلاك 3 أزواج من المجموعات الصبغية، ما يعني 6 مجموعات من الصبغيات. لأجل هذا يمكن القول بأنها كانت نباتات سُداسيّة المجموعات الصبغية hexaploides، وتعني hexa ستّة وتعني ploide مجموعة صبغية. لكن يملك البطّيخ الأحمر نوع Citrullus lanatus 11 زوج من الصبغيات فقط { أي أنه لا يكون من مضاعفات العدد 3 بالمُطلق }. بالتالي لماذا يؤكِّد العلماء حصول التضاعف الثلاثيّ الغريب؟ بمجرّد تحقيق سَلْسَلَة جينوم البطّيخ الأحمر وتحديد المناطق ذات التضاعف الثلاثيّ، كانت تلك المناطق خاضعة للترتيب. بيَّن ذاك الترتيب 7 مقاطع متضاعفة ثلاثيا كبرى وهو ذات عدد المقاطع الملحوظة في الدالية { شجرة العنب }. بالنسبة للعلماء يُمثِّل كل مقطع متضاعف ثلاثياً كروموزوم قديم، هكذا يطرحون النموذج التالي: سلف يمتلك 7 أزواج كروموزومات أنجب سلف يمتلك 21 زوج كروموزومات { الكروموزوم = صبغي }. وقد تطورت من هذا السلف المُمتلك 21 زوج كروموزومات الورديّات. لكن وعلى اعتبار أن الكروموزومات لا تشكّل كيانات مستقرّة، فقد كان بالإمكان حصول اندماجات وانشطارات { فواصل } فيها لأجل خلق الجينوم الراهن الذي يمكننا رؤيته اليوم في البطيخ الأحمر. لهذا لا يبدو جينوم البطيخ الأحمر سداسيّ المجموعات الصبغية hexaploide بفضل اعادة الصياغة المتكرِّر والمسمى paleohexaploide والذي يعني " مجموعة قديمة بستة كروموزومات أو بستة مجموعات صبغية ". لكن الأمر لا يتوقّف عند هذا الحدّ فقد تسلَّل إلى جينوم البطيخ الأحمر الكثير من الينقولاتtransposones. والينقول عنصر جينيّ يملك القدرة على الدخول لوحده في الجينوم، والقدرة على الخروج من منطقة الجينوم المندمج فيها ليتسلَّل لمنطقة أخرى أو حتى التضاعف دون إذن. تمثل هذه العفاريت الجينية { الينقولات } ما نسبته 42.5% من حجم الجينوم .  تطوّر جينوم البطيخ الأحمر: 1 جينوم مُضاعف الصيغة الصبغية سلف. 2 جينوم سداسي الصيغة الصبغية سلف. 3 إعادة تنظيم جينيّ. 4 غزو الينقولات الكبير. 5 جينوم بطيخ أحمر حديث البطّيخ الأحمر وأصله اللغز لكن المفارقة تكمن باكتشاف الماضي البدائيّ البعيد للبطّيخ الأحمر، في حين يبقى ماضيه القريب لغز. على الرغم من وجود العديد من الفرضيّات، كالتي تقول بأنّ بطيخنا الأحمر المزروع ربما قد أتى من الحنظلCitrullus colocynthis L. Schard وموطنه شمال أفريقيا. لكن هذه الفرضيّة لا تكون قويّة كفاية راهناً، يشكّ العلماء اليوم بأنّ السلف الأرجح لبطيخنا الأحمر المزروع هو تنوعات Citrullus lanatus المسماة Citrullus lanatus var. Citroides والمعروفة باسم citrón أو tsamma. ولأجل البحث عن أسلاف في عالم النبات، بالغالب يتم اللجوء لجينوم البلاستيدات الخضراء cloroplastos وهي أعضاء الخليّة المسؤولة عن التركيب الضوئيّ ولا تكون أكثر من بكتريا قديمة متعايشة أو متكافلة. تكون الفكرة المركزية بسيطة: عندما يتشابه هذا الجينوم عند بلاستيدات نباتين أكثر، فهذا يعني أنهما أكثر قرابة من بعضهما البعض. لكن هذا الأمر لا يكون بسيطاً في حالة البطيخ الأحمر. على مستوى جينوم البلاستيدات الخضراء، كان البطيخ الأحمر المزروع var. Lanatus مُختلفاً عن البطيخ الأحمر البري var. Citroides، بينما وبذات الوقت كانت الانواع المختلفة من var. Lanatus { البطيخ الأحمر المزروع } المدروسة غير ممكن التمييز بينها. وحتى لم يتمكنوا من إيجاد فروقات بين سلالة var. Lanatus المسماة بالبطيخ Egusi وسلالات أخرى من var. Lanatus مختلفة المزايا التكوينية والتغذوية. الأمر الذي لفت الانتباه هو أنّ نوع البطيخ Egusi { الذي يسمى كذلك bara } هو نوع تقليدي في بلدان كنيجيريا أو الكونغو، حيث تستمر زراعته بقصد انتاج البذور { الغنيّة جداً بالبروتينات والكاربوهيدرات } والتي ستشكل مكون مألوف في النظام الغذائي لسكّأن تلك المنطقة. أما الثمار فغير قابلة للأكل { ففيها لبّ أبيض قاسي ومرّ } حيث يمكن استخدامها كعلف للماشية. اذاً هو نوع مختلف جذرياً عن البطيخ الأحمر التقليدي الذي نشتريه من السوق. بالعودة لعلم الوراثة / الجينات، فلقد استخلصت دراسة لاحقة مؤسَّسة على تسلسلات ومقارنة جينات بلاستيدات خضراء عديدة { مأخوذة من بطيخ أحمر مزروع بكل أرجاء العالم } شيء مشابه لما سبق ذكره أعلاه. مع ذلك يلاحظ الباحثون بأنّ النوع var. Citroides كما النوع var. Lanatus يكونا مرتبطان بصورة وثيقة مع النوع Citrullus ecirrhosus { المتوطّن في صحراء ناميبيا }. لهذا اقترحوا فرضيّة تقول، بأنّ كلا تنوعيّ Citrullus lanatus أي var. Lanatus و var. Citroides ربما قد تطورا بصورة مستقلة عن النوع Citrullus ecirrhosus، وبناء على هذا سيكون هذا النوع هو النوع السلف لكلا التنوعين. لكن تبقى تلك الفرضية عبارة عن مخاطرة تقتضي دراسات جديدة للبرهنة عليها.  إعادة بناء الشجرة النشوئيّة التطوريّة لأنواع مختلفة ومتنوعة من الجنس Citrullus ما الذي يقوله البحث المنشور هذا العام في مجلة الطبيعة Nature والمبني على سلسلة جينوم البطيخ الأحمر؟ يقول أشياء هامة جداً، حيث أنه يتفق مع الأعمال التي أشرنا لها سابقاً ويحقق نقلة نوعيّة. بدايةً، هو لا يتحدث عن تنوعات بل عن أنواع فرعيّة subespecies { بالرغم من امكان استخدام " التنوعات " المذكورة أعلاه }. وبحسب هذا البحث فإنّ النوع الفرعيّ { لبطيخنا الأحمر } vulgaris يكون، إثر تحليلات مركّزة، غير قابل للتمييز عن النوع الفرعيّ mucosospermus { البطيخ الأحمر Egusi }. وهذا ينسجم مع ما رأيناه آنفاً، فقد كان البطيخ الأحمر Egusi على مستوى جينوم البلاستيدات الخضراء, غير قابل للتمييز عن أعضاء آخرين ينتمون للنوع var. Lanatus { بطيخنا الأحمر ذاته }. كذلك نلاحظ في بحث مجلة الطبيعة بأنّ النوع الفرعيّ mucosospermus يكون ذو ارتباط وثيق بالنوع الفرعيّ lanatus وسيحصل ذات الأمر من ارتباط وثيق بين var. Lanatus وبين var. Citroides. ما يكون أكثر أهميّة في بحث مجلة الطبيعة هو محاولة إعادة بناء الشجرة النشوئيّة التطوريّة للنوع Citrullus lanatus، ولهذا قاموا بتحقيق السلسلة وإجراء المقارنة بين المادة الجينية للنواة الخليوية لأنواع بريّة فرعية وصيغ مزروعة من نوع البطيخ الأحمر هذا. وكنتيجة فقد لاحظوا الارتباط التالي، سيتولد من النوع الفرعيّ subespecie lanatus { المُكافيء للنوع var. Citroides } النوع الفرعيّ subespecie mucosospermun { أي البطيخ الأحمر Egusi } ويمكن أن يكون هذا الأخير هو السلف المُباشر لبطيخنا الأحمر المزروع والمسمى بالنوع الفرعي subespecie vulgaris. كذلك تؤكّد الدراسة الأخيرة بأنّه وبالرغم من التنوُّع الهائل مورفولوجياً في مختلف أنواع البطيخ الاحمر، تكون الفروقات الجينية ضئيلة على الأقلّ بالمقارنة مع أنواع مزروعة أخرى منه. ومن المؤسف عدم إدخال النوع C. ecirrhosus في الدراسة لتأكيد أو نفي دوره بهذا التاريخ للبطيخ الأحمر.  درجة القرابة بين أنواع بطيخ Citrullus lanatus مختلفة. يمثّل اللون الوردي النوع subsp. Lanatus، واللون الأزرق النوع subsp. Mucosospermun، اللون الأخضر النوع subsp. Vulgaris المزروع في أميركا واللون الأحمر النوع subsp. Vulgaris المزروع في شرق آسيا إعادة بناء تدجين البطيخ الأحمر بما نمتلك من معلومات, يمكننا البدء في فهم ماضي بطيخنا الأحمر وكيف أمكن للتدجين التأثير فيه. لكن وكما هو معتاد فكل تأريخ يمتلك بداية وتلك البداية تبدأ مع البطيخ البرّي. على الرغم من امتلاك هذا السيناريو لمشكلة، حيث أنّ البطّيخ البرّي يتقاسم مساحة توزّعه الطبيعي مع أنواع مزروعة وأنواع مزروعة قد أضحت بريّة وكذلك إمكان حصول تهجين بينها وبين أنواع مزروعة أو عائدة لتكون بريّة. مع هذا سنحاول تقديم صورة هي الأقرب للوضع الأصليّ للبطيخ الأحمر sandía. يمكن أن يشكِّل نوع البطيخ La sandía tsamma الشهير باسم Citrullus lanatus تنوُّعاً ضمن citroides أو مثل Citrullus lanatus كنوع فرعيّ من lanatus بحسب الكاتب، فكما رأينا أعلاه، يمكن أن يكون المُكافيء الأقرب للإصدار البرّي والأصليّ للبطيخ الأحمر sandía. ينمو نوع البطيخ البرّي tsamma بصورة طبيعية بمناطق عديدة من القارة الأفريقية بما فيها صحراء كالهاري. يكون متكيفاً بصورة ممتازة مع المناخ الجاف والحارّ. ينبت إثر انتهاء فترة الإمطار مباشرة { بين شهري يناير / كانون ثاني وفبراير / شباط } ويكتمل نموّه ليصل البلوغ بأواسط فترة الجفاف { شهري أبريل / نيسان ومايو / أيار }. تكون ثماره كبيرة نسبياً مقارنة " بثمار نبات بري تقليدي "، ويمكن أن يبلغ وزن البطيخة الواحدة إلى 776 ± 358 غرام ومحتوى مياه يصل إلى 89% من وزنها. يكون مثيراً، فما الذي تريده نبتة صحراوية من إنتاج ثمار ضخمة هكذا ومليئة بالمياه بحقبة الجفاف؟ يبدو أنه مضيعة للوقت! سيما حين ننتبه لحال نبات الصبَّار الذي يسعى جاهداً لتوفير حتى قطرة مياه واحدة. لكن من المؤكّد أنكم قد خمَّنتم سبب تلك الاستراتيجية. نثر البذور. نجد في مناطق أوروبا الرطبة الكثير من النباتات التي توفِّر فواكه ملفتة للانتباه ومليئة بالسكريات لعدد من الطيور والثدييات { كحالة توت العلِّيق }. في الصحراء يحظى توفير المياه للكائنات الحية الأفضلية كما أوضح هذا فيلم رانجو. تُبرز دراسات حقلية استهلاك تلك الفاكهة { البطيخ البري بصحراء كالهاري } من قبل قوارض صغيرة، والتي تقوم إثر تحطيم القشرة القاسية الخارجية بفتح الباب أمام حيوانات صغيرة أخرى وفقاريات. لكن ربما يشكّل هذا مشكلة وتقوم تلك الحيوانات بالتسبب بأذيّة أكثر من تقديم فوائد: يقومون بأكل بذور البطيخ كونها كبيرة ومغذية. ولهذا تقوم تلك القوارض بالاحتفاظ ببعض تلك البذور في مخازنها وينسون بعضها بوقت لاحق كما يحدث مع السناجب والبلُّوط أو الجوز. وربما بفضل تلك القوارض الصغيرة قد طوَّر البطيخ قشرة صلبة ليتم فتحه ويُستهلك من قبل ثدييات كبيرة. وهذا ما تمت ملاحظته في نوع البطيخ sandías tsamma والمُستهلَكْ من قبل عواشب { حيوانات آكلة للأعشاب } ولواحم { حيوانات آكلة لحوم } كبيرة، يمكن للضبع نوع Hyaena brunnea تناول 18 بطيخة بليلة واحدة، بينما يمكن لنوع الغزال Oryx gazella استهلاك 5 إلى 6 بطيخات يوميا لتلبية احتياجاته من المياه { وهذا متاح حيث يمكن للنبتة إنتاج 2 إلى 4 بطيخات بالمتر المربّع الواحد }. ولهذا تحوّلأ نوع البطيخ sandías tsamma لبديل للحصول على المياه خلال حقبة قاسية جافة بالصحراء, بينما تحصل الثدييات الكبيرة على المياه والاغذية منه، وتناثر البطيخ وتوفُّر بذوره عبر فضلات الحيوانات أينما تنقلت.  مشهد من صحراء كالهاري: بطيخة نوع sandías tsamma /Citrullus lanatus var. citroides silvestre برّي ويظهر غزال يتناول طعامه في حقل بطيخ منه يعلِّمنا علم البيئة أسباب امتلاك البطيخ الأحمر لتلك الملامح المميِّزة له: ثمرة تنحو لتكون كبيرة { جداً }، قشرة قاسية نسبياً ولبّ مليء بالمياه. وبصورة شبيهة بكون هذا مفيداً للحياة البريّة ولا يكون غريباً أن يكتشف البشر مصدر للمياه والمواد المغذية كهذا. تعود أقدم الوثائق الخاصة باستخدام هذا النبات لموقع Ti-n-Torha Two Caves في منطقة Uan Muhuggiag بالصحراء الليبية. وقد سكنت هذه المنطقة ومنذ العصر الحجري قبائل تمارس الصيد والتقاط الثمار، وتشتهر منطقة Uan Muhuggiag برسومها القديمة في الكهوف والتي تعكس مشاهد الصيد والرعي. هناك عثروا على أوائل المؤشرات على استعمال البطيخ الأحمر. وقد عثروا على بذرة من نوع بطيخ Citrullus colocynthis على الأقلّ وكذلك بذرة لنوع بطيخ Citrullus lanatus بطبقات تعود لما قبل 9000 عام من خلال استخدام الكربون 14، لكن يفضل الباحثون أن يكونوا حذرين لدرجة يتم بها تجاهل هذه اللقية خشية أن تكون البذور حديثة ووصلت لطبقات أرضية قديمة. مع هذا وفي ذات الموقع قد عثروا على بذور عديدة من نوع البطيخ Citrullus lanatus والتي تعود لما قبل 5000 عام، وُجِدَتْ تلك البذور إلى جانب بقايا نباتات وأعشاب بريّة أخرى، بينما تنتمي بقايا حيوانات متوفرة بذات المكان لحيوانات داجنة { ماعز وأغنام } والتي تعكس وجود مجتمع رعاة. ربما يُشير حضور تلك البذور لأن الناس التي سكنت المنطقة، ربما قد استخدمت هذا النبات كأعلاف للماشية، وربما كغذاء { سواء من ثمرته أو بذوره }، لكن لا يوجد إشارة لبدء زراعته.  في اليسار، رسوم بكهف Uan Muhuggiag، في اليمين، بذور معثور عليها في الكهف ذاته: a وb منظور خارجي ومقطع طولي بالبذرة لنوع بطيخ Citrullus colocynthis، c وd منظور خارجي ومقطع طولي لبذرة نوع Citrullus lanatus بالوقت الراهن وفي تلك المناطق الحارّة، للآن يمكن العثور على غنى كبير بأنواع البطيخ المزروعة، بما فيها ما يعتبره الباحثون سلالات باكرة للنوع المزروع كالنوع الحاضر في صحراء كالهاري. تقترح الدراسات الجينية من جانبها بأنّ منطقة جنوب القارة الأفريقية { كيب تاون، ناميبيا وجنوب أفريقيا } ربما تشكِّل المنطقة التي بدأت فيها زراعة نوع البطيخ sandía tsamma وأعطت تلك الزراعات " البدائيّة " لهذا النوع بدورها نوعنا الراهن من البطيخ الأحمر var. Lanatus المُنتشر في كل أرجاء العالم حالياً. مع ذلك يرافق علم الآثار عملية تدجين النوع Citrullus lanatus بمناطق واقعة إلى الشمال وبالتحديد في مملكة مصر العليا القديمة وخلال حقبة السلالة الخامسة { بين 4000 إلى 3000 قبل الميلاد }، واعتمد الآثاريون على فاكهة وبذور تنتمي لهذا النبات وعُثِرَ عليها في القبور العائدة لتلك الحقبة. اضافة لتقرير مهم آخر عن بقايا نوع البطيخ Citrullus تعود لما قبل 1330 عام قبل الميلاد وفي قبر الفرعون توت عنخ آمون بالتحديد. كذلك ظهرت بقايا لهذا النبات منتمية لفترة ما قبل 1500 عام قبل الميلاد في السودان. زرع المصريون القدماء هذا النبات، كما قامت بهذا شعوب أفريقية أخرى. بوقت لاحق انتشرت زراعة البطيخ الأحمر بقسم كبير من حوض البحر المتوسط وخلال الألف الأوّل قبل الميلاد وقد وصل أوج زراعته خلال الحقبة الرومانيّة.  سلالات بطيخ حاضرة في موزمبيق: a وb سلالات مًنتجة للبذور، c سلالة يجب طبخها قبل استهلاكها، d وf سلالات يمكن تناولها نيئة وتشكل " بطيخ الصحراء الحلو " الشهير لكن تدجين هذا النبات كان له ثمن. فعلى سبيل المثال، يمكننا قراءة دراسات مُحققة منذ أكثر من عقد، حيث يتم فيها إجراء مقارنات بين سلالات بطيخ مختلفة قاطنة في ناميبيا. وهناك يمكن العثور نوع بطيخ tsamma برّي وأنواع مزروعة أخرى كالسلالات التجارية التي يمكن العثور عليها في السوق,،سلالات محلية ببلدات ناميبيا ويمكن تناولها نيئة، سلالات بطيخ مزروعة يمكن طهي لبها كي تصبح قابلة للأكل ولإنتاج بذور يستهلكها الإنسان. في هذه الدراسة تمت المقارنة بين ملامح عديدة: الشكل, الحجم، اللون، عدد وبنية البذور، الثمار، الأوراق.. الخ بكل سلالة بطيخ حاضرة. لا غرابة بكل هذا التنوُّع. فعلى مدار أجيال وأجيال، زرعت البلدات الأفريقية هذا النبات لاستخدامه بمجالات شتّى. ففي موزمبيق تتم زراعة البطيخ بمناطق زراعة البقول { الذرة بقسمها الأكبر أو السورغم }. ويساهم اختلاف أنواعه بتوفير استخدامات مختلفة. يتم استهلاك الثمار بصورة طازجة، يمكن تحضير مشروبات غير متخمرة أو تحضير نوع من النبيذ المسمى xicalavatla، كما أنه يمكن طبخ الأوراق الفتيّة والثمار للاستهلاك البشريّ، استهلاك البذور بصورة طبيعية أو محمصة ويمكن تحضير الطحين منها ويُستخدم كتوابل. كما أنّ سلالات أخرى تُستخدم كعلف للحيوانات. سمحت دراسة بطيخ ناميبيا بتكوين شجرة نشوء تطوريّة لهذا النبات، للسلالات المزروعة لانتاج البذور وتلك السلالات المخصصة ثمارها للطبخ المُسبق، وذلك لانها كانت ذات ارتباط مع الصيغ البريّة للنبات. وربما لهذا السبب يمكن اعتبارها " زراعات بدائيّة "، أي النباتات الأقرب لأوائل السلالات التي ظهرت إثر البدء بتدجين هذا النبات. من جانبها، تشابهت السلالات التي أمكن تناولها طازجة { نيئة } مع السلالات التجارية المُنتَجَة على مستوى عالمي. وربما قد جرى انتقاء تلك المزايا التي جعلت بالامكان استهلاك تلك السلالات بصورة نيئة، وهي التي أعطت الأصل للسلالات المزروعة الحالية في العالم. وفي ذات الدراسة، لا يُلاحَظ وجود توافق بين المنطقة الجغرافية والشبه، وقد أرجع الباحثون هذا لتاريخ البطيخ في تلك المنطقة الحافل بالاختلاط بين سلالات برية وأخرى مزروعة محلية.  ترتيب سلالات متنوعة للبطيخ الحاضرة في ناميبيا، وذلك بحسب مظهرها / شكلها: W نوع برّي، S نوع منتج بذور، C نوع يجب طبخه، L سلالة محلية تُستهلك نيئة و M سلالة تجارية عالميّة وتستهلك نيئة بيَّنت نتائج البحث المحقّق في ناميبيا على البطيخ الاحمر البري sandía tsamma وعلى السلالات المحلية المزروعة، بأنّ التدجين قد أدَّى لتحقيق سلسلة من التغيرات الهامة: نموّ بحجم الثمرة { فالأنواع المزروع ذات ثمار أكبر بكثير من الانواع البرية }، بالتوازي مع هذا يكون حجم البذور عند تلك السلالات المزروعة أكبر بالرغم من تناقص عددها. كذلك قاد التدجين لحدوث نموّ بمعدّل الإنبات، حيث يكون معدّل الإنبات بالسلالات البرية مُنخفض وغير منتظم، بينما يكون معدّل الإنبات في الأنواع المزروعة عالي وموحّد. لكن ربما تكون النتيجة الأهم لتدجين البطيخ هي القضاء على الطعم الغير مستحبّ فيه. فأنواع البطيخ البري بما فيها أنواع Citrullus ecirrhosus و Citrullus colocynthis كما نوع البطيخ sandía tsamma الذي ينمو كأيّ عشبة بريّة في أماكن عدّة بأفريقيا: يحتوي لبُّها على مادة اسمها curbitacina تساهم بخلق الطعم المرّ الزائد عن حدّه. تُشير الدراسات الجينيّة / الوراثيّة لأن الطعم المرّ في البطيخ قد جرى تعديله من خلال جين محدّد يسمى suppressor of bitterness. في هذا الوضع، تمتلك النباتات البرية للإصدار السائد من الجين السابق. بينما تمتلك النباتات المزروعة حلوة الطعم لاصدار متنحي من ذاك الجين فقط. تمّ الحصول على هذا الاصدار المتنحي من الجين بفضل طفرة بسيطة ساهمت بتعطيل الجين المسؤول عن انتاج الطعم المرّ. وبالواقع يمكن العثور على نوع البطيخ البري sandías tsamma بطعم غير مرّ ببعض الأحيان وبعض الأماكن. ولهذا لن يكون غريباً أن يقوم المزارعون بانتقاء وزراعة تلك النباتات غير المرّة.  سلالات بطيخ متنوعة. نوع فرعيّ subsp. Lanatus الموافق للنوع sandía Tsamma، النوع الفرعيّ la subsp. Mucosospermun الموافق للنوع sandía Egusi، النوع الفرعيّ subsp. Vulgaris وتشكل كلها أنواع تجارية بجنوب شرق آسيا وفي أميركا يتميّز البطيخ كذلك بلون اللبّ. ففي الأنواع البرية منه { Citrullus ecirrhosus, Citrullus colocynthis و sandía tsamma silvestre } لا يكون الطعم مرّ فقط بل يكون لون اللبّ أبيض حتى. مع ذل, يمكن أن يكون لبّ الأنواع المزروعة أحمر اللون، برتقاليّ، على لون سمك السلمون، أصفر. وهذا يعود بحسب ما علمنا علم الوراثة لتأثير عدد من الجينات المسؤولة عن تنظيم اللون. لكن تكون الأمور معقّدة حتى الآن. ففي كل نوع بطيخ، يوجد اصدارات عديدة للجين ذاته. سنضع مثال على ذلك، ففي أحد أنواع البطيخ يمتلك " الجين المسؤول عن لون البذور " اصدارين: {1} إصدار يُنتج بذور صفراء اللون و{2} إصدار آخر يُنتج بذور بلون أخضر. اذا بسطنا الأمر، يمكن القول بأنّ النباتات { ونحن } نمتلك جميعنا وكل واحد من جيناتنا يمتلك نسختين، بمعنى لدينا " نسختان " لكل جين. نرث " نسخة " من أمنا و"نسخة " أخرى من أبونا. يمكن أن تكونا " النسختين " متكافئتان: ذات الاصدار لهذا الجين. على سبيل المثال، فيما لو ترث نبتة البطيخ " جين مسؤول عن لون البذور " بإصدار أصفر وإصدار آخر أصفر، فلن يكون هناك أيّ شكّ بأن النبات سيقوم بإنتاج بذور صفراء فقط. وسيتعقد الأمر فيما لو تمتلك كل " نسخة " لإصدار مختلف بذات الجين. ففي حالة كتلك، غالبا ما يكون إصدار للجين أكثر " قوّة " من الإصدار الآخر له، بكلمات أخرى: يسود إصدار للجين على الإصدار الآخر. وفي مثالنا السابق، فيما لو يسود إصدار الجين المسؤول عن اللون الأصفر على الإصدار المسؤول عن اللون الأخضر فستورث النبتة لأجل " الجين المسؤول عن لون البذور " إصدار أصفر وآخر أخضر، وستنتج النبتة بذور بلون أصفر بحسب درجة سيادتها.  قواعد الوراثة. يمثّل A الإصدار السائد الأصفر لإعطاء لون البذور. a يمثِّل الإصدار المتنحي الأخضر لاعطاء لون البذور. أما أحرف اليسار فتمثل التوريث الأموميّ، بينما أحرف اليمين فتمثل التوريث الأبويّ يحصل في البطيخ أمر فيه الكثير من الشبه. بوجود أكثر من جين مسؤول عن تنظيم عمليّة تكوين اللون. وكذلك يمتلك كل جين أكثر من نُسخة / إصدار لنشاطه الخاص ذاته، بصيغة يمكن من خلالها تمييز الجينات التالية: - White flesh: يسود الإصدار Wf ويكون مسؤول عن اللون الأبيض بلبّ البطيخ ويكون الإصدار wf متنحي، لكن سيكون هذا الإصدار مُعطِّل للجين. يكون هذا الجين مثيراً، حيث أن حضور الإصدار Wf يقضي بظهور آثار جينات أخرى ليس لها أيّ تأثير على اللون. - Yellow flesh: يكون الإصدار Y سائد وهو المسؤول عن اللون الأصفر في اللبّ، أما الاصدار y فيكون متنحي ويعطي اللون الأحمر، وباعتباره متنحي وفي حال حضور الإصداران سويّاً Y و y فلا آثار تُذكر. بمعرفة هذا ورؤية المثال السابق الخاص بلون البذور، سنعرف بأنّ " النسختين " لكل جين بذات النبات يمكن أن تشكِّلا ذات الإصدار للجين أو يمكن أن يشكِّلا نسختين مختلفتين. ووفق هذه الصيغة وبحسب النسق الجيني سيكون لدينا لون الفاكهة متغيِّر وفق الآتي: - WfWf YY أو Wfwf YY: لبّ أبيض اللون، لدينا إصدار سائد للجين Wf وهو المسؤول عن ظهور آثار الجين Y الغير قويّة بصرياً. - wfwf YY أو wfwf Yy: لبّ أصفر اللون، لدينا إصدار مُعطّل من الجين wf لحضور إصدار سائد أقلّ من الجين Y والذي أمامه لا يمكن للإصدار المتنحي y أن يفعل شيئاً. - wfwf yy: لبّ أحمر اللون، لدينا إصدار معطّل من الجين wf ولا وجود لأيّ إصدار سائد للجين Y ولهذا سيظهر الإصدار المتنحي من الجين y. قد يبدو سهل وبسيط، لكن الواقع لا يكون بكل تلك البساطة. ويُضاف لما ورد: إصدار آخر للجين y هو المسمى y0 المسؤول عن انتاج اللون البرتقالي { لون لحم سمك السلمون } جين اسمه Canary Yellow flesh وعبر نسخته السائدة C يحقق اللون الأصفر الليموني، وجين يسمى inhibitor of canary yellow والذي يقوم بنسخته المتنحية بإيقاف عمل الإصدار / النسخة C. فيما لو يكن هذا كافيا حتى الآن، يوجد حالة كاكاو شبيهة من حيث لون البذور، لون قشرة الثمرة، شكل الثمرة وملامح أخرى. ما نتعلمه من هذه الحالة هو أن كل الكائنات الحيّة تمتلك قدرة كامنة هائلة للتغيير بفضل التنوُّع بالمادة الوراثيّة / الجينيّة الذي تمتلكه تلك الكائنات، وكذلك أنّ قوة انتقائيّة، كالتدجين أو الانتقاء الطبيعي يمكنها تعديل الكثير بالكائن الحيّ بناء على هذه القاعدة.  لون لبّ البطيخ بحسب عمل الجينات كما رأينا سابقاً: Wf لأجل لبّ أبيض، Y لأجل لبّ أصفر والمتغيّر المتنحي y لأجل اللبّ الأحمر. وهنا نشير لأنّ متغيرات الجينات تسمى أليلات. وتسمى الأليلات الغير سائدة بالأليلات المتنحية فلقد أمكننا رؤية ما الذي فعله التدجين مع البطيخ من تعديلات، وسنتابع تاريخه من اللحظة التي وصلنا لها أعلاه. فقد انتشر البطيخ من أفريقيا باتجاه الهند { بحدود العام 800 ميلادي } والصين { بحدود العام 1100 ميلادي }، ثم لاحقا امتدت زراعته لكامل قارة آسيا خلال القرن الخامس عشر. ويعتبر باحثون آخرون بأنّ حضور البطيخ يعود لفترة أبكر في الجنوب الشرقي الآسيوي، أي في الهند، نيبال، بورما، تايلند والباكستان باعتبار هذه المنطقة شكّلت مركزاً لتدجين البطيخ. الجنوب الأفريقي، وسط القارة الأفريقية والجنوب الشرقي الآسيوي: يُجمع كثير من الباحثين على اعتبارها نقطة الانطلاق بعمليّة التدجين للبطيخ. مع ذلك لا يتعارض هذا مع اقتراح حصول تدجين مستقلّ بمناطق مختلفة من العالم، علماً أنّ أحدث الدراسات بهذا الصدد تقترح بأنّ القارة الأفريقية تشكِّل مهد البطيخ المُدجّن والمكان الذي انتشرت منه زراعته بمناطق أخرى من العالم. وبهذه الصيغة تقترح الدراسات الجينيّة بكون أنواع البطيخ المزروع في الهند، أوروبا والولايات المتحدة الأميركية: تجد أصلها في البطيخ المزروع في جنوب أفريقيا. وربما يشكِّل أول إدخال للبطيخ لأوروبا، هو ما جلبه العرب معهم منه لإسبانيا، كما تُشير بعض السجلات في قرطبة { العام 961 ميلادي } وبإشبيلية { العام 1158 ميلادي }. ويتفق هذا مع ما ذكرناه في مقدمة هذا البحث، حيث أنّ sandía تأتي من العربيّة الفصحى " سنديّة – نسبة لبلاد السند "، بينما يسمونه في إقليم كاتالونيا بطيخ الجزائر meló d’Alger أي يرتبط بالقارة الأفريقيّة. كانت عملية انتقال زراعة البطيخ لشمال أوروبا بطيئة جداً، وفي الواقع لم تصل زراعته لبريطانيا حتى بدايات القرن السادس عشر ميلادي، وربما يلعب مناخ تلك المناطق الدور الأكبر ومدى ملاءمته لمزروعات أفريقية، ولهذا كان يتوقف انتشار تلك المزروعات وفق إيقاع تكيفها مع المناخ الجديد. بالنهاية وفي القرنين السادس عشر والسابع عشر، سافر البطيخ سواء مع مستعمرين أوروبيين أو مع عبيد أفارقة بصورة مُباشرة من أفريقيا باتجاه شمال القارة الأميركية وجنوبها، حيث انتعشت زراعته وتجارته جداً هناك.  يتبع يتبع |
|
|
|
|
|
|
08-11-2016, 06:12 PM
|
رقم الموضوع : [10] |
|
عضو ذهبي
|
البُستان التطوريّ {10}: تطور الموز يكون الموز { Banana للموز المُستورد لإسبانيا، وPlátano للموز البلدي الإسباني } عبارة عن ثمرة سهلة الحمل، حيث يتميز سطحها بخاصية عدم الانزلاق ما يساعد على الإمساك بها، يُشير لونها لحالة النضج من عدمه، فحين تظهر البقع السوداء على سطحه يكون بلغ وقت النضج وبالتالي وقت الاستهلاك المثاليّ. يسمح طرف الموزة بفتحها بسهولة. يكون قشر الموز قابل للتحلل بسهولة وتناول اللبّ يسير على الإنسان. كما أنّ هذه الثمرة سهلة الهضم { ولا تمتلك بذور }. كل هذا لا يمكن أن يكون صدفةن بالتالي الله موجود!!! وثمرة الموز هي أكبر كابوس يمكن للمُلحد أن يواجهه!! أليس هذا غير قابل للدحض؟ بالنسبة لي لا أفهم، بكل صراحة، كيف تكون الأشياء " كاملة " بالنسبة للكائن البشريّ؟. هل فاجأكم هذا البرهان الذي قدّمه الخلقيّان Kirk Cameron و Ray Comfort منذ بضع سنوات، ألا تصدّق هذا؟ أتستغرب وصول الخلقيين لهذا الدرك؟ ويمكن رؤية هذا الفيديو للتأكُّد من هذا. لن أراهن على حصان خاسر، لا أقول بأنّ الموز لا يكون مثالي للاستهلاك البشريّ، لكن ثمار أخرى لا تكون هكذا ببساطة. لنذكر الجوز وثماره وما يجب القيام به لاستهلاك لبّه أو تين الصبّار وأشواكه اللاسعة وبذوره الداخلية المزعجة للكثيرين أو غيرها الكثير. سنبدأ بهذا الموضوع بتقصّي أصول الموز البلدي والمُستورد واللذان بقيا هكذا لأننا نحن رغبنا به.  الكثير من أنواع الموز في العالم قبل المتابعة يجب التنويه لأننا حاولنا تحصيل كل التفاصيل الهامّة لتقديم رؤية شاملة ومُختصرة قدر الإمكان لأصل وتطوّر وتدجين الموز، لم يكن الأمر يسيراً جداً فالمعلومات متوزعة على عدد هائل من المقالات والمراجع العلمية. ولهذا يجب تقديم الشكر لتسهيل دخولنا إلى المجلة المرموقة PNAS Proceedings of the National Academy of Sciences of the United States of America والتي شكّلت معطياتها العمود الفقري بهذا الموضوع. يمكن القول أننا أمام نصّ محضّر من عدد كبير من الاخصائيين علمياً والعاملين في مراكز أبحاث مُشتركة بين استراليا، بلجيكا، الولايات المتحدة الاميركية، فرنسا، Guadalupe و Vanuatu، كما أنه مصدر مفتوح يمكن لمن يود الاطلاع رؤية الروابط التالية: •Perrier, X. et al. 2011. Multidisciplinary perspectives on banana (Musa spp.) domestication. PNAS 108 (28): 11311-11318. مفاهيم مُسبقة لأوّل مرّة سنحاول، بسلسلة البستان التطوريّ، تقديم موجز مُسبق. نرى أنه من المفيد توضيح مصطلحات سيكثر استعمالها خلال المقال، وذلك لأجل الوصول لفهم موضوع الموز ذاته وما الذي حصل به حتى وصوله لصورته الراهنة، كالآتي: - الجينوم: هو كامل المادة الجينية بالكائن الحيّ. يتكوّن الجينوم من كل الجينات وكل جين بكائن حيّ، كما يحتوي على أشياء " ليست جينات " لكنها ايضاً مصنوعة من الحمض النووي الريبي منقوص الاوكسجين DNA. - الكروموزوم { الصبغي }: لا يكون الجينوم هو ذاته في النباتات والحيوانات، بل يجري تقسيمه لقطع بأطوال مختلفة، يسمى كل مقطع من تلك القطع بالكروموزوم أو الصبغي. - المجموعة الصبغية: يمتلك كل نوع حيّ نباتي أو حيواني عدد محدّد من الكروموزومات { الصبغيات } باحجام وأشكال مختلفة فيما بينها. ستكون ploidía " المجموعة الصبغية الأساسيّة ". تمتلك الخلايا الجنسية { كالبويضة، الحيوان المنوي، حبوب الطلع } مجموعة وحيدة من الصبغيات ولهذا تسمى haploides { وحيدة المجموعة الصبغية }. بصورة عامة تمتلك خلايا الجسم الأخرى مجموعة مضاعفة من الصبغيات ولهذا تسمى diploides { ثنائيّة المجموعة الصبغية }. تظهر ببعض الحالات كائنات لديها ثلاث مجموعات صبغية بخلاياها وتسمى تلك الكائنات triploides { ثلاثيّة المجموعات الصبغية }.  مجموعات صبغية أحادية المجموعة الصبغية، ثنائيّة المجموعة الصبغية وثلاثية المجموعة الصبغية من اليسار إلى اليمين بالترتيب. القسم الأوّل: العالم اليوم / من اقتصاد الكفاف إلى اقتصاد الرأسمال تكون ثمار الموز، في أوروبا وأميركا الشمالية، ذات شعبية كاسحة. لكن تكتسب في بلدان أخرى أهمية أكبر نظراً لامتلاكها قيمة حيوية. يشكّل الموز المُنتج الغذائي الرابع الأكثر أهميّة في العالم وتكون نسبة استهلاكه 85% في مناطق مدارية وتحت مدارية ويكون الموز الغذاء الرئيسيّ لحوالي 400 مليون شخص. يعني هو الفاكهة الأكثر انتاجاً بمستوى عالمي كالأرز، القمح والذرة، ويُعتبر أحد أهم الزراعات الغذائيّة في العالم. يوجد أكثر من 1000 نوع مزروع من الموز، وتمثل عملية إنتاج سنوية بحدود 105 إلى 120 مليون طنّ متوزعة على 150 بلد. مع ذلك تكون ما نسبته 90% من هذا الانتاج مخصصة للاستهلاك المحلي في بلدان، مثل: أوغندا، راواندا، الكاميرون أو الغابون حيث تبلغ نسبة استهلاك الفرد من الموز سنويا حوالي 100 إلى 200 كغ. استفادت بلدان أخرى من تجارة الموز، سيما بلدان مثل الإكوادور، كولومبيا، كوستاريكا، غواتيمالا والفيلبين. يكون أهم مصدر منتج ومصدّر للموز في أوروبا: جزر الكانارياس { إسبانيا }، حيث أنتجت في العام 2009 حوالي 352 ألف طنّ من موز الكانارياس الشهير، نعم إننا ملكية موزية بامتياز!!  موزنا الناجح في جزر كانارياس، تنوعات قادمة من Cavendish مع هذا وعلى الرغم من وجود أكثر من 1000 نوع موز، فإنّ أكثر من نصف الإنتاج العالميّ لمختلف الأنواع آتية من تنوعات Cavendish وبإنتاج سنوي يصل لحدود 60 إلى 65 مليون طن. كذلك لدينا الموز المخصص للطبخ بأفريقيا وآسيا وبإنتاج سنوي يصل لحدود 45 إلى 50 مليون طن، وتتمثل خاصيته الرئيسية باحتياجه للطبخ ليسهل تناوله. ويتحرك بكلا نوعي الموز رأس مال قدره حوالي 7900 مليون دولار سنوياً. تكتسب تنوعات Cavendish من الموز أهميّة كبرى، وفي الغالب يعرف غالبيتنا أحد متحدريها. فعلى سبيل المثال، يمتلك موز جزر كانارياس تنوعات هي Giant Cavendish، Dwarf Cavendish وسلالة جديدة مولودة بذات جزر الكانارياس هي Gruesa Palmera بين تنوعات أخرى. تشكّل تنوعات مثالية بمناخ تلك الجزر، حيث تنمو بكثافة كبيرة بحقول الزراعة، تقاوم الأمراض الخطيرة على الموز وسهلة القطاف نسبياً. تكون طبيعة ثمار تنوعات Cavendish من الموز ومتحدريها معروفة جيداً: لبّ لحميّ، نسيج ناعم وطعم شبيه بحلوى شهيّة، كما أنها لا تحتوي على بذور. تشكّل ثمار غنيّة بالطاقة { غنيّة بالكاربوهيدرات }، كما انها تحتوي على كميات كبيرة من الفيتامين ب6 وكميّة أكثر من مقبولة من الفيتامين سي. بالرغم من اعتبار الموز فاكهة مثالية عند كثير من الاشخاص، فإنّ تاريخها يُبرز غرابة بالنوع الأصل الذي تطورت منه. فالموز البرّي السلف لأنواعنا المزروعة اليوم: يكون عبارة عن ثمرة مختلفة كثيراً وصعبة الهضم، وكما قال الحكيم كونفوشيوس:" ربما تساوي صورة واحدة: 1000 كلمة "!.  إلى اليسار ثمرة موز مزروع تنوعات Cavendish مألوفة بأسواقنا. إلى اليمين ثمرة موز بريّة Musa acuminata malaccensis نوع Pahang أحد أسلاف الموز Cavendish، هل رأيتم البذور؟!! القسم الثاني: عائلة من إلاهات الإلهام الجميلات / شعر علميّ ينتمي الموز لفصيلة Musaceae التي يعود أصلها لمناطق استوائيّة بأفريقيا، آسيا واستراليا. تتميّز نباتاته بالضخامة وبأوراق خضراء جميلة، لا يكون شجراً ولا شجيرات، في الواقع يكون عشباً قد بلغ حجماً ضخماً. ويرى أخصائيو علم النبات حتى الآن وجود ثلاث أجناس بهذا النبات، هي: Musa ، Ensete و Musella. يمكن العثور على الجنس Ensete بمناطق مدارية بأفريقيا وجنوب شرق آسيا، ويكون وضعه قيد النقاش حتى الآن، حيث يوجد بحسب دراسات علمية عدّة من 5 إلى 9 أنواع تنتمي لهذا الجنس. تشكّل أعشاب ضخمة يمكن اسخدامها كغذاء، مصدر للألياف النباتيّة ولها قيمة تزيينية بكثير من البلدان. تُشبه ثماره ثمار الموز المألوفة، لكن حجمها صغير، جافة، مليئة بالبذور وغير قابلة للأكل، اضافة لأنّ النبتة الأم تموت إثر إعطائها الثمار. يشمل الجنس Musella نوع واحد فقط هو Musella lasiocarpa وهو نبات قيِّم أصله يعود لجنوب شرق الصين وفيتنام يُعرف بنطاق اخصائيي الحدائق باسم " زهرة اللوتوس الذهبية " أو " الموز القزم الصيني ". يمتلك بالوقت الراهن قيمة تزيينية كبرى بمجال الحدائق، وقد جرى نقله لأماكن عديدة في العالم.  إلى اليسار نموذج من موز Ensete glaucum، إلى اليمين نموذج من موز Musella lasiocarpa لكن ما يبرز بين الموزيات هو الجنس Musa والذي جرى وصف أكثر من 50 نوع ينتمي له، كما ينتمي له الموز الغنيّ الذي نأكله كلنا، يعود أصله للجنوب الشرقي الآسيوي بجزره وأوقيانوسيا. ومن زمن ليس ببعيد تعرّف أخصائيو النبات على الموز المزروع حامل الاسم Musa paradisiaca وهو ذات الاسم الذي وضعه عالم الطبيعة السويدي الشهير كارل لينيو Carl von Linneo في العام 1753. يوجد الكثير من التأمُّل بالاسباب التي دفعت كارل لينيو لاختيار اسم Musa لهذه الفئة النباتية، فهل كان هذا الاختيار لأنهم أسموه musa في مصر وأماكن أفريقية أخرى في القرن الخامس عشر ميلادي كتشويه للمصطلح العربي " موز mouz، maouz، moz أو maws " والتي ترمز لمدينة Moka؟. وأخشى أن أقع بالخطأ باسم المدينة، فربما تكون هي المدينة الحالية التي اسمها المخا Mocha أو Mokha الواقعة في اليمن. أو ربما يكون كارل لينيو قد اختار الاسم تيمناً باسم عالم النبات وطبيب الامبراطور أغسطس { القرن الأوّل قبل الميلاد } وكان اسمه Antonio Musa؟ أو ربما محاكاة لإلهامات inspiradoras musas بإلوهيّات الفنّ والعلوم في أساطير اليونان والرومان؟ مهما يكن السبب: فإنّ هذا الاسم يستحضر الشعر!! مع هذا فقد تغيرت كثير من الأمور وتُعلِّمُنا الاكتشافات الجديدة بعلم النبات وعلم الوراثة بأنّ كارل لينيو قد كان على خطأ. يكون وضع النوع Musa paradisiaca قيد النقاش وما استخدمه كارل لينيو كنموذج للموز المزروع في الواقع كان عبارة عن نوع هجين من نوعين. أيّ نوعين كانا؟ حسناً هذه قصّة تبدأ في مزيج من الجزر، الأرخبيلات والصخور القائمة بجنوب شرق آسيا وأوقيانوسيا.  نماذج موز Musa x paradisiaca لغايات تزيينية وهي تقريبا النبتة التي ألهمت كارل لينيو القسم الثالث: مولود في الأحراج / شهادة للقتل إثر عقود من الدراسات، يوجد اليوم اتفاق كبير حول أصول وأسلاف الموز الذي نشتريه من الأسواق بالعادة. ويُعتبر أهمهم النوع Musa acuminata Colla الذي يعود أصله لجزر جنوب شرق آسيا، أندونيسيا، ميكرونيزيا وميلانيزيا، والأهم جزيرة غينيا الجديدة. يعتبر أخصائيُّو علم النبات النوع البرّي Musa acuminata " كعشب غابة " ونبات متشرِّد! تلعب هذه العشبة في النظام البيئيّ الغابي دوراً تعبوياً، فعندما تفرغ مساحة ما من الغابة وتحت أي ظرف، سينمو فيها النوع البري Musa acuminata ويغطيها، سيكون هذا عبارة عن غزو مؤقّت، فدوماً هناك نباتات أقوى ومُقاومة أكثر والتي يمكنها أن تحلّ مكان نباتات أخرى، وحتى وقتها يستفيد الموز البري ويستغلّ زمن سيطرته لأقصى حدّ ممكن. ستُزهر أشجار الموز البرّي إثر نموها وستكون أزهارها محطّ اهتمام واستفادة عند عدد من الفقاريات بغية تحصيل النكتار { الرحيق } مثل نوع الخفّاش Cynopterus sphinx وطيور من نوع Arachnothera magna ونوع Arachnothera longirostra. وخلال سعيهم لتحصيل الرحيق يساهمون بنشر حبات الطلع بين الأزهار الذكرية والأنثوية وتحقيق الإلقاح والإخصاب.  إلى اليسار زهرة جميلة للنوع Musa acuminata مع طائر Arachnothera magna. إلى اليمين خفّاش Cynopterus sphinx يضع قدميه في زهرة النوع Musa sp يمتلك هذا الجهد بالإخصاب سبباً هاماً هو زيادة التنوع الجيني / الوراثي للبذور وبهذا تزداد احتمالات البقاء على قيد الحياة. مع هذا فلا يكون كافياً لتوفير مستقبل لائق للمتحدرين { للنسل }. فستحتاج البذور للانتشار لأماكن أخرى بهدف امتلاك حظوظ أوفر للإنتاش والنموّ وذلك بالعثور على مكان ما مفتوح وسطة عتمة وكثافة الحرش المداري. لهذا تُنتج نباتات الموز البرّي كميات كبيرة من الفاكهة على مدار العام. حيث تعطي تلك الثمار ألياف تحظى بتقدير نوع الخفاش Cynopterus sphinx الذي سيقطفها ويأخذها بعيداً لمكان يمكنه اكلها فيه بهدوء. كما أنّ هذه الثمار جاذبة لزباد النخيل الآسيوي في جزر لاس بالماس Paradoxurus hermaphroditus والذي يأكلها بشراهة وحين يطرح برازه سيساهم بنقل البذار لامكنة أخرى من الغابة وهي اسمدة طبيعية بكل حال. لهذا تكون أهمية هذه الحيوانات بالغة لنبات الموز ونثر بذوره أي تكاثره. تكون تلك البذور كبيرة الحجم نسبياً، ولهذا تشكّل وجبة دسمة للقوارض من امثال السنجاب Callosciurus erythraeus، السنجاب Callosciurus caniceps، السنجاب Menetes berdmorei، الجرذان Rattus sp والفئران Mus sp، دون نسيان حيوان ثديي آخر ليس بقارض رغم شبهه به: هو الزباب Tupaia glis.  إلى اليسار ثمار موز برية Musa acuminata، لاحظوا البذور الكثيرة. إلى اليمين زبّاد Paradoxurus hermaphroditus كناشر طبيعي لبذور هذا النبات جرى التفكير سابقاً بأنّه حين يتمكن خفّاش من تناول الموز البري فربما نحن كبشر يمكننا تناوله ايضاً. لكن هذا يشكِّل رأي متسرّع. صحيح أن الموز يؤكل من قبل البشر نيئاً ومطبوخاً، وحتى يوظفوه كعلاج للاسهال والزحار، لكن حذار! فثمار الموز البريّة Musa acuminata وأنواع أخرى لا تكون صحيّة، ففي الواقع يصل تناولها ليشكّل خطراً على الصحّة!! وهذا لا يشكل مزحة، فاستهلاكها قد يؤدي لحصول مشاكل تصل حدود الإنسداد المعويّ الخطير جرّاء تراكم fitobezoares. وما هي fitobezoares؟ هي تراكمات المادة النباتيّة غير القابلة للهضم. وتنتج هذه التراكمات بحالة تناول الموز البري وذلك لاحتوائه على مركبّات مسماة taninos أو pectinas ستتحوّل في الجهاز الهضمي لنوع من الاسمنت المتحد والمندمج بألياف عديدة وبذور ضخمة للثمرة وبهذا يتم خلق كرة قادرة على سدّ المعي.  بازهرات { تراكمات مادة نباتية } ناتجة عن استهلاك ثمار الموز البري Musa acuminata. وجرى أخذها من ذات المريض وهي بقطر 5 سنتمتر ويمكنها إعاقة عمل الأمعاء وحال حصول هذا مع أي شخص فلن يتم إنقاذه إلا عبر عمل جراحيّ. وقد اشتكى كثير من الأطباء في جزيرة Laos من ظهور حالات خطيرة جرّاء استهلاك الموز البري، وفي الواقع لا يستهلك سكّان لاوس هذا الموز إلا جرّاء الفقر ونقص الموارد بتلك الجزيرة. وإذا اتبعنا منطق الخلقيان Kirk Cameron و Ray Comfort يمكننا الاستنتاج بأنّ الله قد وهبنا ثمرة مميّزة ينتهي تناولها بعمل جراحي!! مع هذا فسيكون من الخطأ اعتبار النوع البري Musa acuminata باعتباره " واحد أو حصريّ ". فكما تكلمنا سابقاً، لقد استوطن هذا النوع بنجاح في عدّة جزر بجنوب شرق آسيا وبأوقيانوسيا، ولهذا تكون جماعات النوع Musa acuminata متناثرة ومنعزلة بكثير من الأماكن. لكن لهذا الإنعزال تبعات فقد راكمت تلك الجماعات، بصورة مستقلة عن بعضها البعض، تغيرات كثيرة على مدار آلاف الأعوام، فهي تكون الآن مختلفة فيما بينها بمرحلة متوسطة بين نوع أم ونوع جديد. وتستقبل هذه الجماعات التي تكون بحال انتقاليّة اسم " نُويع أو تحت نوع subespecie ".  نشوء الأنواع جرّاء الانعزال الجغرافيّ ويسمى بالاسبانيّ Especiación alopátrica ويكون حضور هذه النُويعات هاماً، فلقد لعبت دوراً مركزياً بأصل وتطوُّر الموز المُدجّن. كما أنها تشكّل بحد ذاتها مثال ممتاز للتطوُّر: حيث ميّز أخصائيُّو علم النبات بين 4 جماعات للنوع البري Musa acuminata بناء على المظهر وشكل الساق، الأزهار، الثمار وملامح أخرى، كذلك تختلف تلك الجماعات الأربعة فيما بينها بناء على الدراسات الجينية المحققة على جينات النواة، الميتاكوندريا والبلاستيدات الخضراء. واللافت هنا هو اتفاق تلك الاختلافات مع اختلاف المناطق الجغرافيّة التي تنشأ بها تلك الجماعات من الموز البري. يتناسب كل هذا مع منحى تطوريّ، حيث أنّ الموز وإثر نشوئه واستمراره منعزلاً في الجزر خلال آلاف أو مئات آلاف الأعوام، قد راكمت جماعاته فروقات كافية فيما بينها لاعتبارها نويعات، جماعات نباتية بطريقها لتصير أنواع مختلفة، تكون تلك النويعات / تحت نوع subespecies هي: 1- النُويع banksii موطنه جزيرة غينيا الجديدة. 2- النُويع malaccensis موطنه شبه جزيرة مالايا. 3- النُويع ثلاثي النُسخ burmannica ، burmannicoides و siamea وهي غير مختلفة جينياً، موطنها تايلند، ميانمار، بنغلاديش، شمال شرق الهند وجنوب الصين، تشبه كثيراً النُويع malaccensis وتتقاسم مناطق حدودية معه ولهذا ربما قد حصلت عملية تهجين بينهما. 4- النُويع zebrina، موطنه جزيرة جافا. يمتلك الكم الأكبر من المادة الجينية للجماعة { حتى 10% أكثر } وربما يكون " السلف " الأقدم. مع ذلك تبيّن دراسات جينية أخرى وجود جماعات بمناطق جغرافية أخرى مختلفة بصورة كافية ليتم تسميتها نُويعات ومنها: - النُويع microcarpa، يحضر بجزيرة بورنيو ويكون قريباً من النُويع zebrina. - النُويع truncata، متوطّن حصرياً بالأراضي المرتفعة بشبه جزيرة مالايا. - النُويع errans، الحاضر في جزر الفيلبين وذو ترتيب تصنيفي حديث.  خارطة التوزُّع الجغرافي للأنواع الفرعية المختلفة للموز Musa acuminata القسم الرابع: ماضٍ بعيد / فترة شباب فوضوية يمكننا البدء بتقصي مسألة تدجين هذه الثمرة، هذا التطور الذي يحققه البشر فيها، فلقد تغيرت الثمرة جذرياً من فاكهة شيطانيّة إلى فاكهة مُباركة. لكن الموضوع أعقد من هذا وأبعد من عملية التدجين ذاتها. بحيث تُعنى هذه السلسلة " البستان التطوريّ " بتقديم ما يكشفه علم الوراثة لماضي النبات المدروس الهام. نحن نكون في عصر الجينوم، وفي السنوات الأخيرة، كرّست مجموعات بحث وقتها لفك شيفرة جينوم كثير من الانواع الحيّة، وجرى التركيز على تلك الأنواع ذات الأهميّة الحيوية الطبيّة، الاقتصاديّة أو التطوريّة. ويمتلك الموز بعض من تلك القضايا، وفعلاً درسه أخصائيُّو علم الوراثة ونشروا بشهر أغسطس / آب 2012 بمجلة الطبيعة Nature بحثاً شارك به 19 مركز بحث من استراليا، البرازيل، الولايات المتحدة، فرنسا، هولندا، تشيكوسلوفاكيا وسويسره وحمل البحث الاسم: D’Hont, A. et al. 2012. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants . Nature 488: 213-217. Artículo de acceso libre. يبيّن البحث أوّل مسودة لكامل جينوم الموز، الموز البري Musa acuminata النُويع malaccensis tipo ‘DH Pahang كأحد أسلاف النوع التجاري الشهير Cavendish. يتكون جينومها بحسب هذا البحث من 11 زوج من الكروموزومات / الصبغيات { أنواع موز أخرى لديها 10 أزواج كروموزومات } و523 مليون زوج قواعد، وتشمل كامل المادة الجينية على حوالي 30000 و40000 منطقة محددة كجينات { نحن البشر لدينا ما بين 20000 و21000 جين } ويحتوي نصف الجينوم، على الأقلّ، على جينات قافزة " الينقولات transposones " وهي مواد وراثيّة مصنوعة من الحمض النووي الريبي منقوص الاوكسجين DNA لكنها أحياناً تغيِّر أماكنها ضمن الجينوم وتتضاعف دون إذن مُسبق، وهي للآن كيانات خاضعة للتدراسة والتمحيص. لكن المعلومة الأفضل قد أتت من تحليل تفصيلي للجينوم. وبدراسة استفادت من كل الدراسات السابقة، بما نشرته مجلة الطبيعة، حيث نرى التركيز على التضاعفات الكاملة للجينوم وآثارها على التطوّر عند سلالات عديدة للموز. يشكّل تضاعف الجينوم الكامل ظاهرة تسببها طفرة وتؤدي لظهور كائنات جديدة بكامل جينومها المتضاعف نسبة لأبويها. لا يشكّل هذا ظاهرة غريبة ويحدث بصورة مستمرة حتى يومنا هذا ويشكّل مصدراً لخلق أنواع جديدة وهذا رأيناه سابقاً في موضوعي تطور العنب والبطيخ.  يرى العلماء بهذه الحالة بأنّه منذ حوالي 65 مليون عام قد حصل حادثي تضاعف جيني بما سيشكل سلف الفصيلة الموزية musáceas. يعني هذا أن التضاعف الجيني المزدوج قد ولّد نوع جديد من النبات وهو هذا سيشكّل بدوره، إثر مرور ملايين الأعوام، سلف الموز الراهن بين أنواع أخرى، وتسمى عملية التضاعف الجيني المزدوج تلك α/β. لكن لا تنتهي القصة هنا، فيبدو أنّ ذاك السلف بدوره للفصيلة الموزية قد ورث تضاعف جيني آخر. بحسب المُشاركين بذاك البحث، فقد حصل التضاعف الجيني الآخر بحدود ما قبل 100 مليون عام وقد تمت تسميته التضاعف γ. ولّد هذا التضاعف سلف رتبة الزنجبيليات Zingiberales الراهنة وهي مجموعة نباتية تتكوّن من فصيلتين شقيقتين هما الموزيات musáceas و الزنجبيليات zingiberáceas وهذه الأخيرة فصيلة نباتية ينتمي لها أنواع مثل الزنجبيل Zingiber officinale. كذلك عثروا على حوادث تضاعف جيني في الفصيلة النجيلية gramíneas { التي ينتمي لها الأرُّز والقمح }، لكن يبدو بأنّ تلك التضاعفات قد حصلت بمعزل عمّا حصل بالزنجبيليات.  إلى اليسار تضاعفات جينومية كاملة حاصلة عند سلف Zingiberales { تضاعف γ } وعند سلف Musa { تضاعف α/β }. إلى اليمين تطور أحاديات الفلقة وثنائيات الفلقة، تمثلا اللوحتان حوادث تضاعف جينومي كامل القسم الخامس:" انظري أماه, فلقد ابتكرت الزراعة " لكي نعرف العلم جيداً سيكون من المفيد معرفة أدواته. ونجد بعلم الآثار النباتيّ واحدة من أهم الأدوات وهي البلورات الحجرية fitolitos، وهي بنى مجهرية تتركّب من ثنائي أوكسيد السيليكون أو السيليكا sílice ويتم تصنيعها داخل الخلايا النباتيّة. ويشكّل هذا المركّب " مادة زجاجية بالغة القسوة " ويمكن حفظها خلال حقب زمنية طويلة. وعلى اعتبار أنّ كل نوع أو جنس نباتيّ يمتلك بلوراته الحجرية الخاصة به، فسيكون بالامكان التحديد بدقّة لزمن تواجد تلك البلورات في الموقع المدروس. تكون بلورات الموز الحجرية شديدة التميُّز لكنها لا تسمح بالتفريق بين الأنواع البرية والمزروعة، ولهذا وعلى الرغم من اثباتها حضور نباتات الموز بمكان ما وبزمن ما، فسيتوجب اثبات التدجين من خلال معطيات أخرى. ربما يعود التاريخ المشترك للموز والبشر لزمن قديم يتوافق مع استيطان جزيرة غينيا الجديدة { منذ حوالي 40000 عام }. يوجد في غينيا الجديدة الموقع الأثري Kuk Swamp على ارتفاع 1560 متر عن سطح البحر ويقع ضمن الحرش المداري بجبال منخفضة وبدرجة حرارة متوسطة هي 19 مئوية ومعدل سقوط مطر هو 27000 ميليمتر سنوياً، نعم هو مكان رطب بالعموم. يكتسب موقع Kuk Swamp أهميته مما عُثِرَ عليه فيه من لقى تعود لما قبل 10220 – 9910 عام، من قبيل: 1- يبيّن سجّل غبار الطلع والبلورات الحجرية لتلك الحقبة حلول أماكن فارغة بموضع الحرش. 2- تظهر ثقوب مكافئة لما يُصنع منها لحمل بعض المزروعات وتدعيم بعضها الآخر. 3- يوجد بقايا سدّ قديم لتجميع المياه ودعم قناة ريّ وعدم استنزاف الارض لها. 4- وفي حال عدم كفاية كل ذلك، فيوجد أدوات حجرية وبقايا نبتة معروفة باسم Colocasia esculenta وهي تشبه نبتة الموز { دون أن تكون نبتة موز } وتستعمل كغذاء. يُشير كل هذا لأخصائيي علم الآثار لبدايات باكرة بأنشطة زراعية بمناطق المستنقعات.  إلى اليسار بلورات حجرية للنوع Musa عرضة لمسح مجهر الكتروني { ما خلا الصورة اليمنى العليا }، إلى اليمين صورة نبات القلقاس Colocasia esculenta حيث يكون ساقه تحت أرضي متضخم وقابل للأكل ستتابع تلك التقنية تطورها في موقع Kuk Swamp. ففي حقبة زمنية تمتد ما بين 6950 – 5990 عام تظهر الثقوب، بقايا الكاربون اضافة إلى تقنية زراعية جديدة مثالية لمناطق رطبة: تلال مرتفعة تسمح بتصريف اسهل للمياه. ربما يكون ما هو أكثر أهميّة هو توفُّر كميات ضخمة من البلورات الحجرية للموز والذي يمكن تفسيره بكثافة زراعة الموز لدرجة لا يضاهيه أيّ نبات يمكن العثور عليه بصورة طبيعية. لهذا يقدِّر علماء الآثار حصول زراعة الموز بفترة ما قبل 6 إلى 7 آلاف عام. كذلك تكون هذه البلورات الحجرية شديدة الشبه ببلورات الموز Musa acuminata النوع الفرعيّ subespecie banksii وهو النوع الفرعي الوحيد للموز الذي ينمو بصورة طبيعية بتلك الجزيرة. ويكون لهذا مدلول كبير، يتوجب أن تكون زراعة نوع ما، بدقّة، في المكان الذي يعيش فيه: بالتالي لا يمكن تدجين نبات من الصفر أي من نوع غير موجود! ولا يقتصر هذا على الموز فحسب، ففي الواقع كثير من النباتات نشأت في غينيا الجديدة وانتقلت منها لاحقاً، مثل النوع المشار له سابقاً Colocasia sculenta، النوع Dioscorea alata، قصب السكّر Saccharum officinarum والموز النوع الفرعي Musa acuminata subespecie banksii بين غيرها من النباتات. وتكون تقنياتها الزراعية متخصصة بمناطق مستنقعات، وقد توجّب أن يكون هذا الابتكار خاصاً بها، وهو ما يكافيء الابتكار المستقلّ الذي حصل في منطقة الهلال الخصيب أو بمناطق عديدة بأميركا الجنوبية وآسيا قديماً. كما فعل سكّان تلك المناطق بتركيزهم على نباتات محلية غنيّة بالسكريات و / أو البروتينات.  أزهار وثمار الموز البري Musa acuminata النوع الفرعيّ banksii بغينيا الجديدة: A أزهار ذكرية. B ثمرة. C ثمرة مفتوحة لتبدو البذور لكن رأينا أن ثمرة الموز لا تُهضَمْ بسهولة، فلماذا ركّزوا عليها. لماذا تُزرع؟ ربما قد حقّق سكّان ذاك الزمن عمليات تسبق الاستهلاك كالسلق، الشوي، الطبخ والسحق اضافة لخلطه مع مكونات أخرى. ربما توصّل سكّان موقع Kuk Swamp لتحسين القيمة الغذائية لتلك الثمار الأصلية. لكن حتى لو اعتبرنا ثمرة الموز غير مفيدة كلياً، فيوجد أسباب أخرى تجعل من المفيد زراعة الموز، فالبقاء على قيد الحياة يعني الاستفادة من كل الموارد المتاحة بأقصى شكل ممكن. وبناء عليه فسكّان جنوب شرق آسيا والمنطقة الهندية – المالاوية قد استفادوا من نبات الموز بطرق كثيرة: 1- يمكن تناول الأزهار، الأوراق النضرة والأغصان بصورة نيئة أو مطبوخة بصور عديدة. 2- تنفع الأوراق بتحضير أغلفة للمواد الغذائية. 3- يمكن استخراج مواد تساعد بالتنظيف وتحضير الورق. 4- يمكن لبعض اجزاء الاوراق الاسهام بتحضير لفافات تبغ وعناصر أخرى. 5- تمكّن مواطني تلك المناطق من استخدامها بأمور طبية علاجية ودينية طقوسيّة. 6- يكون مفيداً كعلف للخنازير واستخدامات منزلية أخرى. يخلق كل هذا أهميّة خاصة لنباتات الموز في الاقتصادات المحلية المتواضعة. حتى لو أضحت ثمار الموز ثمار عابرة للحدود عالمياً، فأنا أقصد الألياف المُستخرجة من أوراق الموز والتي ساهمت بخلق صناعة نسيجية واعدة، ثياب ذات جودة عالية، حبال، سجّاد، ستائر وأدوات زينة قيِّمة. في الواقع يقترب نوع الموز Musa textiles من نوع مزروع أكثر منه برّي، فهو يشكّل قاعدة " القنِّب " في الفيلبين وهو عبارة عن نسيج نشره الاسبان منذ أواسط القرن الثامن عشر، وشكّل صناعة قويّة جرى تصدير بضاعتها إلى أميركا واليابان، لكن انتاجها قد تدهور بصورة ملحوظة إثر ابتكار الألياف الصناعيّة { بالرغم من دوام استخدامها حتى يومنا هذا }. بدوره ازدهر نوع نسيج آخر مصنوع من ألياف نوع موز برّي آخر هو Musa balbisiana وسوقه في تصاعد بيومنا هذا، وتبلغ الأجور المدفوعة 200000 دولار لتحضير النسيج الذي يُصنع منه اللباس التقليدي الياباني الكيمونو kimono.  القسم السادس: حقبة التغيُّرات والهجائن / حروب الاستنساخ سيبدأ أصل وتطوُّر الموز المدجّن مع أوائل محاولات زراعة النوع Musa acuminata النوع الفرعيّ subespecie banksii في غينيا الجديدة منذ ما يقرب من 7000 عام، وبعد هذا الزمن بفترة بسيطة سيتم إعطاء الخطوة الكبرى التالية بتطور هذا النبات. هاجر سكّان غينيا الجديدة كحال كل البشر بقصد البحث عن مستقبل أفضل وعملوا بالتجارة مع ناس آخرين، هكذا نقلوا فكرة زراعة الموز كما نقلوا الموز المدجّن مسبقاً من النوع الفرعي subespecie banksii إلى جزر أخرى، وكانت تلك الجزر موطناً منذ آلاف الأعوام لأنواع موز فرعية أخرى أصيلة مثل Musa acuminata. سبّب هذا النقل لتلك الأنواع الخصبة لتلك الجزر الغريبة حصول عملية التهجين بين أنواع فرعية محلية. مع هذا ولكونها أنواع فرعية مختلفة فيما بينها، أتساءل لماذا لم يعتبر أخصائيُّو علم النبات بكون تلك النباتات كأنواع مختلفة، فعلى سبيل المثال عندما يتم تهجين فرس وحمار فيعطي هذا التهجين بغلاً وهو حيوان قويّ وذو مزايا جسدية جيدة لكنه يكون مصاب بالعُقم. ويحصل ذات الشيء مع بعض الأنواع الفرعية المختلفة ل Musa acuminata كآبائه، تكون الهجائن عبارة عن نباتات مزدوجة المجموعة الصبغية diploides مع مجموعتان صبغيتان { مجموعتا كروموزومات } { المسماة AA، لكل مجموعة حرف }، حيث تنتمي مجموعة صبغية لنوع فرعيّ وتنتمي المجموعة الثانية لنوع فرعي آخر. يحقق التزويد الجيني / الوراثيّ هذا النموّ الجيّد للنباتات، كما أنه يعطيها مزايا مقبولة من المزارعين. لكن يكون هذا التزويد الجيني ذاته غير متوافق مع نفسه لتوليد خلايا جنسية بحسب الهجين ذاته، حيث لا يتأثّر إنتاج البويضات ولكن سينخفض انتاج حبوب الطلع بنسبة تتراوح بين 50 و 100 %. بناء عليه يمكن تأكيد مسؤولية هجائن كتلك عن انتاج نباتات مصابة بالعقم جزئيا أو كلياً، وبتعبير آخر اعطاء نباتات لا تُنتج بذور. توجّب أن يرافق تلك الظاهرة ميزة أخرى لموزنا المزروع، هي: " الإثمار البكريّ partenocarpia " أي إمكانيّة إنتاج الأزهار الأنثوية للثمار دون حصول الإلقاح. فغالبيّة الأنواع النباتية لا تُنتج الثمار قبل حدوث الإلقاح، فالثمرة بالنهاية هي استثمار طاقيّ ضخم للنبات، يكون الهدف الوحيد منها هو نشر البذور. مع ذلك تنتج الثمار في الموز دون الاحتياج للإلقاح { فكما رأينا، عادة ما تكون الهجائن مصابة بالعقم }، ولكن ما هو سبب هذا؟  جرى نشر أحد أوائل الأبحاث بهذا الاتجاه في العام 1953، وقد عمل عليه عالم النبات الأخصائيّ بعالم الموز Norman Willison Simmonds { 1922 – 2002 } والمؤلّف للعديد من الدراسات مثل " الموز " { العام 1966 } و " تطور الموز " { العام 1962 } اضافة لنشره أكثر من 250 مقال علميّ. وقد اعتبر بذاك البحث بأنّ وجود 3 جينات مرتبطة بالإثمار البكريّ، يكون الجين الأهمّ بينها والحاضر في نباتات مزروعة " P1 " ذو ميزة سائدة، بحيث يكون قادراً على إنتاج ثمار بصورة باكرة { دون إلقاح partenocárpicos }. ويسميا الجينان الآخران " P2 " و " P3 " يمتازان بخاصيّة ثانوية ويظهرا في نباتات بريّة وتكون وظيفتهما فيها مجهولة حتى اللحظة. اقترح الدكتور Dr. Simmonds ببحثه ذاك الفرضيّة التي تعتبر أن هذا الجين قد ظهر جرّاء حدوث طفرات حوّلته من جين بوظائف غير معلومة إلى جين متسبب بحصول الإثمار البكريّ، وبهذا وفي وقت لاحق وعندما فعل الانتقاء فعله عبر عملية التدجين، سادت النسخة " P1 " { المنتجة لنباتات إثمار باكر } عند النباتات المزروعة. ربطت أبحاث لاحقة بين حضور النسخة السائدة لهذا الجين " P1 " بعملية زيادة حجم الثمار fructificación، لاحظوا أنّ الجين السائد " P1 " يمكن أن يترافق جزئياً مع إنتاج ثمار ذات وزن وحجم أكبر { أي ازدياد مردودية النبات }. هل كان البحث عن ثمار أكبر كل مرّة أكثر هو العامل الذي انتقى الإثمار البكريّ بالتوازي؟ أو هل كان البحث عن الإثمار البكريّ هو العامل المتسبب بنموّ الثمار بالحجم؟ أو كلاهما معاً؟ نحتاج لتحقيق المزيد من الأبحاث والدراسات بهذا الإطار. طالت دراسة أخرى النوع البري Musa acuminata النوع الفرعيّ subespecie microcarpa بجزيرة بورنيو { منتج لأزهار أحادية المسكن flores monoicas أي على ذات النبتة يوجد أزهار أنثوية من جانب وأزهار ذكرية من جانب آخر }، جرى درس الإثمار المبكِّر عبر منع وصول المُلقحين للأزهار الأنثوية، وبحسب النتائج فإنّه في حال غياب الإلقاح، ستقوم الأزهار الأنثوية بإعطاء ثمار صغيرة الحجم، والتي سريعا ما تتخرّب فتنفتح وتموت. مع هذا فمن المثير أن جميع نورات النبات قد أعطت ثمرة أو أكثر بعملية مكافئة للإثمار البكريّ أو المبكِّر. تُعتبر ملاحظة وجود أزهار إثمار مبكِّر وغير مبكِّر بذات النورة النباتيّة مثيرة للاهتمام بالنسبة للباحثين، ويكون من تبعاتها اعتبار الإثمار البكريّ خاضع سواء للتحكُّم الجيني / الوراثيّ كما للتحكُّم البيئي.  مجموعة أزهار انثوية للموز Musa acuminata النوع الفرعيّ malaccensis de Borneo. لم تكن الأزهار ملقحة، لكن يمكن ملاحظة أزهار غير مثمرة { لا يوجد نمو للثمرة، فهي تبدو صغيرة بلون أسود وميتة وأزهار مثمرة الثمرة قيد النمو وبلون أخضر. A التفصيل وB عنقود موز برؤية أوسع تكون الميزة الاساسيّة الأخرى في موزنا المزروع، هي: قابليته للأكل. فتناول الموز لا يحمل أيّ تبعات سلبية على الجهاز الهضميّ. يرى العلماء بأنّ هذه الميزة، أي تطوّر اللبّ قد حدث تحت التحكُّم الجيني وتمّ انتقاؤه في الزراعات البدائيّة لهذا النبات، إلى جانب حالة العقم والإثمار البكريّ. ربما ظهرت أوائل العيِّنات القابلة للأكل من الموز البدائيّ بين الموز البرّي Musa acuminata النوع الفرعيّ subespecie banksii بجزيرة ساموا، وبينت تلك العينات أوّل تطوُّر جزئيّ في اللبّ بالرغم من كونها خصبة وبريّة. يرى العلماء بأنّ هذه الخاصية المتمثلة بقابلية جزئية للأكل قد جرى انتقاؤها من النوع الفرعي subespecie banksii ومنه انتقل وانتشر { وربما توسّع } من خلال عمليات التهجين. حسناً فيما لو تكن تلك الهجائن مصابة بالعقم جزئيا أو كليا، فكيف يمكن الوثوق بإنتاج البذور لزراعة مناطق أخرى أو استبدال النباتات التي تموت، كيف يكون بالامكان زراعة هذا النبات؟ يمتلك الموز ورقة رابحة في تكاثره، وهذا يكون من الاسباب التي قادت لتهجينه، فعندما يُنتج حالة العقم، فهذا يمكن أن يقود لتوليد أنواع نباتية جديدة في عالم النبات. تستطيع نباتات الموز إنتاج سلسلة من البراعم الجانبية على الساق والتي من خلالها تنمو " نباتات الموز الأبناء " أو الأفرع، وما هي إلاَّ نُسخ طبق الاصل عن آبائها وتمتلك القدرة على النموّ وصولاً لتحل محل آبائها وأشقائها. مع ذلك، عمد المزراعون بصورة تقليدية إلى فصل تلك الأفرع عن النبات الأم والسعي للحصول على نباتات جديدة منها. وتكون هذه التقنية واحدة من تقنيات عديدة أخرى كاستعمال الجذور والساق الغليظة التي تنمو عبرها الأفرع، والتي تبقى بشروط محددة قابلة لإنتاج أبنائها بصورة أكبر.  تكاثر الموزيات. إلى اليسار برعم موزيّ مولود من بذرته { حيث يشير سهم أبيض }، إلى اليمين فرع أو ساق يؤخذ منه البراعم الجديدة " النُسخ " بناء على هذه التقنيات بزراعة الموز ومتحدريه، تكون كلها عبارة عن اصدارات متطابقة، فالأبناء سيمتلكون ذات مزايا وصفات الآباء، ولهذا نجد في كبرى مزارع الموز نباتات " متطابقة ". بالرغم من أنّ الواقع لا يكون بهذا التبسيط، حيث يحدث بكثير من الأحيان أن يكون المُستنسَخْ ≠ مُتطابق. أهدانا الخيال العلميّ صور جنود مستنسخين متطابقين يسيرون نحو حرب النجوم، تحاول " الحركات المناهضة للاستنساخ " تخويفنا من إعادة خلق هتلر جديد، قام صحفيون وكتاب آخرون بخلق صورة عن مستنسخين متطابقين فيما بينهم كلياً. يكون كل هذا عبارة عن أنصاف حقائق. يحدث بصورة طبيعية أن تتكاثر كثير من الكائنات الحيّة عبر استنساخ ذاتها نفسها. وفعلياً هذا ما يقوم به الموز كتقنية مُستخدمة بزراعته. لكن يحصل عند تلك النباتات ما يسمى " بالطفرات الجسدية mutaciones somáticas " وهي الطفرات التي تحصل في خلايا مسؤولة عن إعطاء البراعم التي تنمو من خلالها الأفرع الجديدة، وكنتيجة لهذا ستكون الأفرع الجديدة عبارة عن نباتات متطفرة ولديها فروقات عن آبائها. يرى أخصائيُّو علم النبات الذين درسوا الموز وتنوعاته الكثيرة، بأنّ تلك الطفرات قد تسببت بنموّ في أنواع الموز عبر الهجائن المختلفة منه. في الواقع وبناء على وجود حالة العقم والإثمار البكريّ، فقد شكَّلت الطفرات الجسدية بالقسم الأكبر من تاريخ تدجين الموز، وتكون المصدر الوحيد للتنوع بهذا النبات، إضافة لظهور خواص ومزايا جديدة فيه. وتُعتبر تنوعات Cavendish مثال جيّد على هذا، وهو مجموعة من الموز المستخدم كحلوى، فقد ساهمت طفرات جسدية بتوليد عدد كبير من تنوعات الموز والتي نجد بينها: Lacatan, Robusta, Valery, Giant Cavendish, Grand Naine, Dwarf Cavendish, Petit Naine, Dwarf Parfitt, Williams و Zelig بين أنواع كثيرة غيرها. وفي وقت قريب، ساهمت طفرة جسدية بالنوع Dwarf Cavendish بإعطاء نوع موز جديد اسمه Gruesa Palmera حصريّ بجزر الكانارياس الاسبانية.  تنوعات موزية: A نوع Red { جينوم AAA } حلو المذاق، B نوع Palayam Codan { جينوم AAB }، C نوع Njalipoovan { جينوم AB } حلو المذاق وخفيف لكن سهل التخريب الذاتي، D نوع Robusta { نوع Cavendis، جينوم AAA } ذو قيمة كبرى بالاستهلاك، E نوع Nendran { جينوم AAB } يستخدم للطهي وعمل تشيبس كما لو كان بطاطا مقلية القسم السابع: أصل نباتيّ / وصول الهجائن مزدوجة الصبغيات diploides من صيغة إلى أخرى، شجّع التهجين ظهور نباتات مصابة بالعقم من جهة، بينما سيشجّع، من جهة أخرى، الإثمار البكريّ { سواء بفعل التهجين أو بتأثير التدجين أو بسبب كليهما } إنتاج ثمار دون بذور بذات النباتات المصابة بالعقم تلك. بذات الوقت سيتم انتقاء ثمار تكون أكبر بالحجم كل مرّة أكثر وذات لبّ أكبر { وربما أمكن للتهجين تقديم العون بهذا الإتجاه }. وتساهم الطفرات الجسدية بنموّ في التنوع الجيني عبر التهجين، ما يؤدي لتوليد تنوّع كبير في الانواع المزروعة منه. لهذا يُعتبر التهجين بين أنواع فرعية مختلفة أحد أهم الخطوات الاساسيّة في تدجين الموز وتحوله من ثمرة مليئة بالبذور وغير قابلة للهضم إلى ثمرة خالية من البذور وسهلة الهضم. ووفق ما تُشير دراسات جينية ولغوية، فقد حدثت أوائل عمليات التهجين عبر " الهجرة من غينيا الجديدة " بحدود ما قبل 6000 عام وهي تتسق مع عملية التدجين التي حصلت قبل 7000 عام للمرّة الأولى. سافر البشر عبر تلك الهجرة من غينيا الجديدة إلى جزر أخرى، وهذا ما ولّد حصول تماس بثلاث مناطق على الأقلّ بين أنواع فرعية للموز: 1- غينيا الجديدة وجنوب جزيرة جاوا. 2- غينيا الجديدة وشرق الفلبين. 3- بين الفلبين وجزيرة بورنيو مع جنوب شرق القارة الآسيوية. ولّدت تلك التهجينات أسلاف كثير من الأنواع المستخدمة حتى الآن من الموز. تكون كل تلك التنوعات مع السلف المُشترك منتمية لذات " المجموعة الفرعية "، ويبرز بينها: 1- Khai و Figue في ماليزيا وجنوب شرق آسيا كنتيجة لتهجين النوعين الفرعيين subespecie malaccensis و subespecies zebrina/microcarpa. 2- Buaya في جزيرة بورنيو كنتيجة لتهجين الأنواع الفرعية zebrina/microcarpa مع subespecies siamea/burmanica. 3- المجموعة الفرعية Galeo في غينيا الجديدة، تكون عبارة عن كوكتيل جيني / وراثي للأنواع الفرعية banksii, zebrina و malaccensis.  مناطق وطرق انتشار المجموعات الفرعية المختلفة للموز الهجين ثنائي المجموعة الصبغية لكن دون شكّ، يستحيل نسيان المجموعة الفرعية Mlali والتي تشمل تنوعات عديدة متوفرة بيومنا هذا فقط في شرق أفريقيا وجزر قريبة { مدغشقر، جزر القمر، زنجبار والجزيرة الخضراء Pemba }. وتُعتبر هذه المجموعة الفرعية قديمة والتي لم تحظَ بكل تلك الشعبية لولا وصولها من مناطق بعيدة، يرى العلماء بأنها نتاج عملية تهجين بين banksii و zebrina/microcarpa وقد حصلت هذه العملية بمكان ما بين جزر جاوا، بورنيو وغينيا الجديدة، مع هذا فلا وجود لهذه المجموعة الفرعية بتلك الأماكن بيومنا هذا. عُثِرَ على أوائل المؤشرات على حضور الموز في القارة الأفريقية بموقع Nkang في الكاميرون. فقد وجدوا بهذا الموقع، بقايا سيراميكية، أدوات حجرية، قطع حديدية، عظام حيوانات، بقايا فحم وثمار جوز متفحمة اضافة لبقايا متفحمة لنوعي الموز Canarium scheweinfurthii و Elaeis guineensis وبفترة زمنية تعود لما قبل 2580 و 2170 عام. وما يهمنا بالموضوع كذلك العثور على بلورات حجرية للموزيات سيما للأنواع Ensete و Musa كما حددهما أخصائيُّو علم النبات. كما يعتبر أخصائيون آخرون أنه يوجد في مواقع مختلفة في Nkang بلورات حجرية للنوع Musa وتعود لما قبل 2790 – 2300 عام. لا يشكّل حضور النوع Ensete أمراً مهماً كثيراً، فمن المألوف العثور عليه في مناطق مدارية بأفريقيا. لكن حضور النوع Ensete يشكّل قصة أخرى، حيث لا تكون هذه النباتات أصيلة في القارة الأفريقية، ويعني تواجدها بأفريقيا أن أحداً ما قد جلبها وزرعها فيها. علما أنه قد جرى تقليديا اعتبار أفريقيا مركزاً مهما لتنوّع نباتات الموز بسبب العدد الهائل من سلالاته وتنوعاته فيها، وقد زُرِعَ خلال قرون بل وآلاف الأعوام. كذلك تدعم المعطيات الجينية والبيوكيميائية فرضية كون القارة الأفريقية مكان حصول تنوُّعات الموز. من حانب آخر، تُشير المعطيات التأريخية واللغوية لأنّ زراعة الموز تعود لما قبل أكثر من 3000 عام. ويقوّي هذا الطرح كل ما عُثِرَ عليه في موقع Nkang. كذلك تثبت لُقى بموقع Munsa الأوغندي لأنّ حضور البلورات الحجرية للجنس Musa يعود لما قبل 4500 عام! وفي حال إثبات هذه المعطيات، فإنّ عملية نقل نبات الموز من جنوب شرق آسيا إلى أفريقيا بهدف زراعته قد حصلت بوقت بالغ القِدَمْ. بالوقت الراهن، يشكّل الموز مصدرا مهما للتغذية في أفريقيا، وقد تولّد عدد كبير من التنوعات بنباتاته بالرغم من لزوم طبخ الكثير منها لاستهلاكه. ويكون من المحتمل أن المصطلح الذي نستخدمه لتسمية الموز banana ذو أصل أفريقي. يشتبه اللغويون بأن المصطلح ما هو إلا امتداد أو تعديل لمصطلحات مستخدمة محليا بأفريقيا لتحديد هذا النبات، مثل «banana» «banaana»، «banna» أو «bana» يسمونه في غينيا bename ومن المثير أنه يوجد بيومنا هذا مجموعة من الموز مسماة «guineos». وتشكل هذه فرضية بديلة لفرضية أقدم تعتبر أنّ أصل كلمة موز banana هو عربي، ويكون المصطلح الأصلي هو البنان ويشبه كلفظ صوتيّ مصطلح banana ويُشير معناه العربيّ للإصبع، ولهذا يرى اللغويون أن العرب استخدموا هذا المصطلح لأنّ كتلة الموز شبيهة باليد وأصابعها وتكون هذه الفرضية عُرضة للنقاش والجدل، فلا وجود لوثيقة عربية قديمة تتحدث عن هذا النبات. بل على العكس من هذا، فمن المعروف بأن اسم الموز قد كان «musa» أو ما يشبهه وقد استخدمه العرب أنفسهم كما رأينا بمقدمة هذا الموضوع.  موز القارة الأفريقية. إلى اليسار تاجر موز ينقل كميّة للبيع قد تصل إلى 30 كغ. إلى اليمين طبق matooke محضّر من موز مغليّ القسم الثامن: مزيد من التعقيد / وصول الهجائن ثلاثيّة المجموعة الصبغية triploides تحدثنا أعلاه عن تهجين أدّى لظهور مجموعات فرعية جديدة من نبات الموز كأسلاف لبعض الأنواع المزروعة الحديثة. سيقوم ذات التهجين بتشجيع ظهور نباتات مصابة بالعقم { دون بذور } وسيجري انتقاء الثمار باكرة الإثمار وذات اللبّ الأكبر لاستهلاكها. كذلك رأينا طرق نشر النباتات المصابة بالعقم، والتي عبر القيام بكثير من " الاستنساخ " للقديم منها لم يتحقق سوى ازدياد أنواع الموز. مع ذلك سيمتلك التهجين تبعات أخرى غير متوقعة من قبل المزارعين. رأينا سابقاً بأنّ التهجين قاد لإنتاج لاتوافق جيني يساهم بقتل خلايا جنسية، الأمر الذي يؤدي لانتاج الاصابة بالعقم. لكن لا يحدث هذا دوماً، فأحياناً يخلق اللاتوافق الجيني خلايا جنسية متطفرة وشاذّة، حيث تمتلك الخلية الجنسية الطبيعية مجموعة صبغية واحدة تسمى haploide، في حين تمتلك تلك الخلايا الجنسية المتطفرة مجموعتان صبغيتان وتسمى diploide سواء كانت بويضة انثوية أو حبوب طلع ذكرية. وهذا ساهم بإنتاج ظاهرة مثيرة للاهتمام. ربما قد ساهمت حبوب طلع ثنائية المجموعة الصبغية diploides بإلقاح بويضات انثوية وحيدة المجموعة الصبغية haploides. أو ربما ساهمت حبوب طلع وحيدة المجموعة الصبغية بإلقاح بويضات أنثوية ثنائية المجموعة الصبغية. تكمن القضيّة بإنتاج هذا الإلقاح لنوع نبات جديد هو نبات ثلاثي المجموعة الصبغية triploide أي يمتلك 3 مجموعات صبغية. وأن يكون الكائن الحيّ ثلاثي المجموعة الصبغية، حيوان أو عند كثير من النباتات، فهذا يعني الوقوع بورطة، حيث تموت الأجنة الناتجة بسرعة هائلة، ولاسباب من بينها أنّ العدد الفرديّ للمجموعات الصبغية يعرقل انقسام الخلايا وكنتيجة يعرقل نمو الجنين. لكن تُشكّل حالة الموز Musa استثناء مذهل، فالأجنة ثلاثية المجموعة الصبغية لا تبقى على قيد الحياة فقط بل تصل لمرحلة البلوغ وبمزايا اضافية. فنباتات الموز هذه المصابة بالعقم وغير المنتجة للبذور، وبفضل مزاياها الجيدة ستجعل البشر مُساهمين بنشرها على امتداد العالم.  يشكّل، هنا، ازدياد عدد المجموعات الصبغية فائدة فورية، تتمثّل بالعبور من نبات ثنائي المجموعة الصبغية إلى نبات ثلاثي المجموعة الصبغية، قد أدّى إلى حدوث نموّ متزامن بوزن وحجم الثمرة، وهو الأمر الذي قاد لولادة نباتات الموز الأكثر إنتاجيّة. يؤكد العلماء بأنّ تلك الظاهرة قد أضحت منتشرة وعامة، وقد حصلت لمرات عديدة في مناطق زراعة الموز في جنوب شرق آسيا والجزر. يشكِّل الموز Mutika / Lujugira واحد من أوائل المجموعات الفرعية ثلاثية المجموعة الصبغية التي ظهرت، ويتواجد بالوقت الراهن في أفريقيا. تسمى هذه المجموعة الفرعية بثلاثية المجموعة الصبغية نمط AAA لامتلاكها ثلاث مجموعات صبغية وكل مجموعة من النمط A { أي ينتمي للنوع Musa acuminata }. تنتج هذه المجموعة الفرعية من عملية تهجين بين النوعين الفرعيين banksii و zebrinaوغالبا قد حصل هذا بين غينيا الجديدة وجزيرة جاوا، على الرغم من غيابها حالياً عن تلك الجزر. من المعتاد استخدام تلك النباتات في الأراضي المرتفعة بالشرق الأفريقي بالطبخ أو لتحضير مشروبات كحولية. مع ذلك، سيدخل لاعب آخر بذاك الزمن إلى المشهد. ويكون اسمه Musa balbisiana Colla وهو نوع بري يعود أصله لمناطق الرياح الموسمية في شمال الجنوب الشرقي الآسيوي، ويتوزع هذا النبات اليوم بجماعات صغيرة ومتفرقة من غينيا الجديدة إلى سيريلانكا. كذلك يمتلك هذا النبات نمط جينيّ genotipo نموذج BB لامتلاكه زوج مجموعات صبغية وكل مجموعة صبغية تكون من النمط B { أي أنها تنتمي للنوع Musa balbisiana }. سيكون هذا التمييز للمجموعات الصبغية مفيداً لنا لمعرفة أصل نباتات موز محددة.  تفاصيل نبات وثمرة النوع Musa balbisiana Colla. A نموذج عنقود لهذا النوع من الموز. B تفاصيل ثمرة ناضجة، لاحظوا كثافة البذور الكبرى. C تفاصيل عنقود ثمار موز غير ناضجة كذلك يكون النوع البري Musa balbisiana Colla مُنتجاً لثمرة صغيرة مليئة ببذور غليظة سوداء، وسيحصل تهجين بينه وبين أنواع فرعية نقيّة وهجينة للنوع Musa acuminata، ما يؤدي لتوليد أنواع موز فرعية جديدة. سيكون أحدها المجموعة الفرعية Popoulou / Maia Maoli وهي مجموعة ناجحة يرتبط اسمها بسلسلة من أنواع الموز المسماة بموز المحيط الهادي، وتمتلك جينوم هو AAB { القسم A ينتمي للنوع Musa acuminata النوع الفرعي subespecie banksii، والقسم B ينتمي للنوع Musa balbisiana }. يشتبه العلماء بأنّ ثمّة زراعات قديمة للنوع Musa balbisiana قد انتقلت من جنوب الصين إلى غينيا الجديدة مجتازة للفيلبين. سيحصل في غينيا الجديدة إعطاء الأصل للمجموعة الفرعية Popoulou / Maia Maoli عبر التهجين ومن هناك ستنتشر زراعته نحو المحيط الهاديء بالتوازي مع الانتشار البشريّ. تكون مجموعة فرعية أخرى هامة هي مجموعة الموز الأفريقي والتي يكون جينومها كذلك AAB { وكالحالة السابقة، يكون الجينوم A عائد للنوع Musa acuminata النوع الفرعي subespecie banksii والجينوم B عائد للنوع Musa balbisiana }. تكون هذه الأنواع من الموز الأفريقي عبارة عن زراعة مستقرة بشكل جيد في المناطق المدارية بأفريقيا الوسطى وفي الغرب منها، بالرغم من أنّ أصلها في الغالب يعود للمنطقة الواقعة بين الفيلبين وغينيا الجديدة. مع هذا لا يزال الطريق الذي سلكوه للوصول لأفريقيا مجهولاً. يبدو أن الاسم المُستخدم ببعض مناطق أفريقيا للموز الأفريقي، يأتي من مُصطلح بلغات البانتو هو kondo، ويُشير هذا لأنّ وصوله لأفريقيا قد حدث قبل انتشار ثقافات البانتو أي منذ حوالي 3000 عام. ويقترب هذا التأريخ من عُمر اللقى المعثور عليها بموقع Nkang { في الكاميرون } البالغ ما قبل 2750 إلى 2100 عام. بالرغم من جهل الطرق التي جعلته ينتشر ويصل لأفريقيا، فإنّ حضور البلورات الحجرية للموز بعمر 4000 عام بمنطقة Kot Diji { في الباكستان } يمكن أن يُشير لأنّ الانتشار قد حصل عن طريق برّي.  مناطق وطرق انتشار مجموعات فرعية مختلفة للموز ثلاثي المجموعات الصبغية القسم التاسع: انتشار عالميّ / على قاعدة النوع المزروع Cavendish بمرور قرون سيحصل حالات تهجين جديدة وحالات ثلاثية المجموعات الصبغية. واحد من أهم تلك الهجائن هو القادم من مجموعتين فرعيتين ثنائيتي المجموعة الصبغية معروفتي الأصل. فسيولد من المجموعة الفرعية Khai الماليزية نوع جديد يسمى Akondro Mainty. وستعطي المجموعة الفرعية Mlali من غينيا الجديدة والجزر القريبة الأصل لنوع جديد يسمى Pisang Pipit. يعتبر العلماء، تدعمهم دراساتهم الجينية ومعرفتهم بأصل وتوزيع الموز تاريخياً، بأنّ الحدث الهام سيحصل بمكان ما بين ماليزيا وتايلند. فهناك سيقوم مشيج أحادي المجموعة الصبغية A للنوع Pisang Pipit بإلقاح مشيج ثنائي المجموعة الصبغية AA للنوع Akondro Mainty. بتلك اللحظة، بالضبط، وُلِدَ النوع Cavendish السلف الرسميّ لموز جزر الكانارياس الاسبانية.  كاريكاتير موزيّ بقالب ديني. صلوا لإلهكم الجديد " موز Cavendish "!! يشكّل موز Cavendish موز ثلاثي المجموعة الصبغية بجينوم AAA وناجح جداً بالوقت الراهن. لكن عندما ظهر هذا النوع، لم يكن وحيداً، فبالكاد منذ 50 عام لم يشكّل زراعة هامة، بل كان شقيقه اللقيط هو من امتلك السيطرة في السوق العالمي. كان اسمه Gros-Michel وهو ثلاثي المجموعة الصبغية ايضاً ومتحدِّر من تهجين Akondro Mainty مع نوع آخر لم يكن معروفاً فيما لو يكن هو Sa أو Khai-Nai-On، يكونا بكل الأحوال كلا النوعين ورثة المجموعة الفرعية Khai. فحتى العام 1950 ، كان النوع المسيطر بالتجارة العالمية هو Gros-Michel ولهذا جرى توسيع زراعته في كثير من بلدان العالم. كما رأينا أعلاه، تتأسّس زراعة الموز على استخدام براعم { فسيلات } النبات، لهذا تكون النباتات الناتجة عبارة عن نُسخ متطابقة مع النبات الأم. تكمن المشكلة الكبرى بأنّ تلك النباتات متطابقة تقريباً بكثير من الملامح بما فيها سهولة التعرُّض لبعض الأمراض. كانت نهاية النوع Gros-Michel على يد آفة مسماة " شرّ بنما " وهي عبارة عن مرض يتسبب به نوع الفطر Fusarium oxysporum Schlect. f. sp. Cúbense الذي يؤثر على الأوعية الناقلة للاغذية والنسغ، ما يؤدي لموت النبات. لم تتمكن كل تنوعات الموز Gros-Michel من عمل أيّ شيء بمواجهة هذا الفطر ما أدى لإنهائها. لا يعرف هذا المرض " شرّ بنما " لا حدود ولا خطوط عرض أو طول، ففي وادي Ulua في هندوراس قد قضى على نباتات الموز بمساحة تتراوح بين 30000 و 40000 هكتار بين العامين 1940 و 1960. لم تنتهِ المشاكل هنا، يمتلك هذا الفطر مقاومة لدرجة ترك المزارعين حقول كاملة دون زراعة لمحاولة قتل الفطر " من الجوع "، وتلك الحقول لم تستخدم خلال 30 عام أو أكثر حتى. كانت الأضرار اللاحقة بالتصنيع وتجارة الموز فادحة ويصعب احصاؤها، وكان المزارعون والمُنتجون على مفترق طرق حاسم: ترك زراعة الموز والوقوع فريسة الجوع أو العثور على مُنقذ يكون قادر على مواجهة " شرّ بنما ". هنا ركَّزوا جهودهم على بعض تنوعات الموز Cavendish، والمفاجأة أنها قاومت الفطر وبالتالي المرض وبقيت على قيد الحياة!!! لاحظ العلماء بأنّ الفطر ينتشر بنوع الموز Gros-Michel دون ضوابط عبر أوعية النبات، مع هذا فبعض أنواع الموز Cavendish تكون قادرة على إفراز مادة جيلاتينية بداخل أوعيتها تعمل على تقييد الفطر وشلّ حركته بداخلها مانعة انتشاره. هكذا يتابع النبات نموّه بظلّ شلّ حركة الفطر تلك بصورة طبيعية. إذاً عثر المزارعون على المُنقذ، ومنذ العام 1960 وحتى يومنا هذا: سيكون الموز السائد على مستوى العالم هو Cavendish.  صورة حقل موز مصاب بمرض " شرّ بنما " في ماليزيا في العام 1995 سيتابع النوع Musa balbisiana حضوره، ولكن تأثيره بالاقتصاد العالمي محدود جداً، حيث يلعب دوراً لدى شعوب تعتمد على الزراعة وذات موارد محدودة واقتصاد كفاف محليّ. سيتشكل في الهند اعتباراً من تهجين نباتات المجموعة الفرعية Mlali مع M. balbisiana المجموعة الفرعية Pome / Prata ذات النمط الجيني AAB، كأقربائه Nendra Padathi و Nadan وهي نباتات موز ذات طعم حلو وحامض بذات الوقت. كذلك ستتشكل نباتات ثلاثية المجموعة الصبغية أخرى من النمط ABB، بالرغم من عدم وضوح أصله بصورة كاملة. في هذه الأنواع من الموز، تأتي مجموعة الكروزومات { الصبغيات } A من النوع banksii وبالتحديد من تنوع قادم من Cavendish. تكون مهمة جداً في شبه القارة الهندية وبأفريقيا. تشكّل هذه الأنواع بعض الأمثلة فقط، فلقد تولّد عن تهجين Musa acuminata و Musa balbisiana عدد هائل من التنوعات وبنسب مختلفة من الجينوم A والجينوم B. كذلك تُشير الدراسات الجينية لأنه عندما ينتج تهجين من هذا النوع، فستكون النتيجة أكبر من مجرّد جمع أجزاء، حيث يتم خلق كائنات جديدة لم تكن موجودة في الطبيعة. ويشكل هذا نتيجة لسلسلة من الظواهر الحاصة جرّاء " مزج " جينومات متباينة بكل هذا القدر: حيث يعيد الجينوم بناء ذاته، عناصر تبدو غير نشطة يتم تنشيطها من جديد، إلغاء جينات أخرى أو تثبيطها، بينما يجري تعديل عمل جينات أخرى، هكذا كما تعمل الجينات الأخرى في شروط أخرى. وفي بعض الحالات، يصل التعديل ليطال بنية الصبغيات ذاتها { الكروموزومات }.  الدورة الثانية للموز ثلاثي المجموعة الصبغية مع أصل وانتشار. تشير الأحرف AAA Cav للنوع Cavendish القسم العاشر: خلاصات / من يستعبد من؟ رأينا تاريخ الموز أعلاه، من كونه ثمرة صغيرة ومليئة بالبذور غير القابلة للأكل عملياً وصولا لثمار لذيذة قابلة للاستهلاك كتنوعات Cavendish. سيتمكن البشر المتخصصون بزراعة وتجارة هذا النبات من تطويع واستعباد نبات يحقق لهم منافع هائلة. وتنوعت أشكال الزراعة من الحقول للبيوت الزجاجية وغيرها. ولكن أشجار الموز فيما لو تمتلك الوعي لذاتها والقدرة على الكلام، ربما لقصّت علينا رواية مختلفة أخرى، هي: هل شاهدتم تاريخ البشر منذ أن عاشوا في قبائل صغيرة وصولاً للمدن الضخمة. لقد تمكنا من استعباد البشر لنشر نوعنا على امتداد العالم واعتباراً من نباتات لا قيمة لها بالاساس. لقد خصّص البشر كثير من الوقت والجهد والمال خلال ملايين السنين للاهتمام بنا والحفاظ علينا، لقد تمكنا من إخضاع الإنسان ككائن يتمتع بالقدرات الأكبر في هذا الكوكب لمصلحتنا.  صورة جويّة لهكتارات مزروعة بالموز في منطقة Innisfail { كوينزلاند، استراليا } استخدم الموز البري في الأحراش حيوانات لتحقيق تخصيب ازهاره ونشر بذوره كالخفافيش والزبَّاد بوصفها وسائط نقل لبذورها. ربما لم تتغير الامور كثيراً في العصر الحديث، فلقد ضحّت نباتات الموز ببذورها لقاء انتاجها لثمار ضخمة، لكن لم يتغير الامر كثيراً حيث يقوم الإنسان العاقل بدور الناقل والناشر لنباتات الموز. وبالنسبة للموز يكون الإنسان أكثر فعالية بهذا المضمار من خفّاش أو زبَّاد!! نحن البشر نتحكم بالنباتات؟ أو النباتات تتحكّم بنا؟ بالواقع لا يشكّل هذا تساؤلاً سهل الإجابة وبتوافق أكبر من بيئة فلسفية ما. ربما يحصلا الشيئين معاً، لكن لا أرى أن كثير من الناس جاهزة للموافقة على أنّ من طُردوا { أبناء آدم } من الجنّة يقبلوا أن يستعبدهم العالم النباتيّ!! وهنا نُنهي موضوعنا الطويل!!  عندما يحكم الموز العالم! عملية قيد الإنجاز يتبع |
|
|
|
|
|
 |
| مواقع النشر (المفضلة) |
| الكلمات الدليلية (Tags) |
| البُستانُ, التطوريُّ |
| الذين يشاهدون محتوى الموضوع الآن : 1 ( الأعضاء 0 والزوار 1) | |
|
|
Powered by vBulletin® Version 3.8.7
Copyright ©2000 - 2025, Jelsoft Enterprises Ltd
diamond










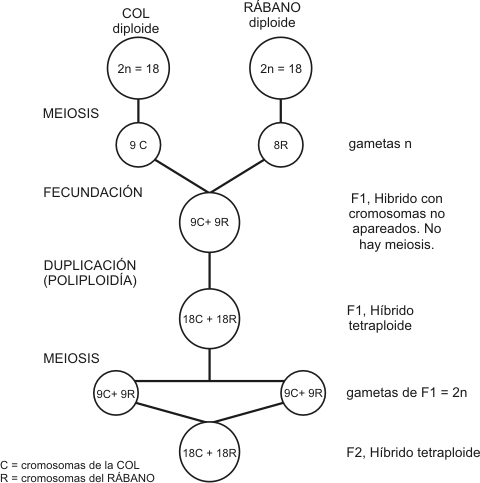







































 العرض العادي
العرض العادي

